Trabajos de investigación
Caracterización molecular del virus de bronquitis infecciosa aviar en granjas de gallinas ponedoras de la provincia de Tungurahua, Ecuador
Molecular characterization of infectious bronchitis virus in laying hen farms located in Tungurahua province, Ecuador
 María del Carmen Revelo-Cueva
Christian Vinicio Vinueza-Burgos
Germán Ernesto Metz
María Gabriela Echeverría gecheverria@fcv.unlp.edu.ar
María del Carmen Revelo-Cueva
Christian Vinicio Vinueza-Burgos
Germán Ernesto Metz
María Gabriela Echeverría gecheverria@fcv.unlp.edu.ar
ANALECTA VETERINARIA
Universidad Nacional de La Plata, Argentina
ISSN: 1514-2590
Periodicidad: Frecuencia continua
vol. 42, e066, 2022
Recepción: 22 Abril 2022
Preprint: 11 Junio 2022
Aprobación: 28 Junio 2022

Resumen: La bronquitis infecciosa aviar genera importantes pérdidas económicas y, si bien la vacunación disminuye dichas mermas, el surgimiento continuo de nuevas cepas virales complica el control de la infección. El objetivo de esta investigación fue caracterizar, mediante técnicas moleculares, las cepas del virus de la bronquitis infecciosa aviar circulantes en explotaciones de gallinas ponedoras de la provincia de Tungurahua, Ecuador. Se trabajó con muestras de hisopados y de órganos de 47 granjas y las secuencias obtenidas fueron comparadas con las correspondientes a 17 vacunas a virus vivo empleadas en la zona. Se encontraron 16 granjas positivas al amplificar la región 5’UTR y un segmento de S1. La construcción de un árbol filogenético mostró que cinco de las secuencias se ubicaron en el mismo clado de las cepas de tipo Massachusetts, linaje GI-1; siete se encontraron en el clado del linaje GI-13 o cepas tipo 793B y cuatro secuencias se agruparon en un clado de cepas tipo Q1, linaje G1-16. Si bien se requieren estudios moleculares más completos en los que se amplifique la totalidad del gen S a fin de obtener datos más concluyentes, se logró determinar que existen tres tipos de cepas circulando en la provincia de Tungurahua, dos posiblemente vacunales y otra de origen desconocido.
Palabras clave: Virus de bronquitis infecciosa aviar, gallinas ponedoras, aislamientos, filogenia, vacunas.
Abstract: Avian infectious bronchitis (IBV) generates significant economic losses. While vaccination decreases these losses, new viral strains are continually emerging. The objective of this research was to perform molecular characterization of IBV strains circulating in laying hen farms in Tungurahua province, Ecuador. Swabs and organ samples from 47 farms were collected and sequenced, and results were compared to 17 live vaccines used in the area Sixteen farms were positive for IBV as determined by amplification of the 5'UTR region and a S1 segment. The construction of a phylogenetic tree showed that 5 samples were in the same clade as the Massachusetts-type strains, linage GI-1; 7 samples were in the GI-13 lineage clade of vaccine type 793B strains, and 4 samples were into a clade of Q1 type strains. In conclusion, although studies that include the entire protein S sequence are necessary, we were able to detect three types of strains circulating in the province of Tungurahua, two of which are probably vaccine-derived and one has an unknown origin.
Keywords: Avian infectious bronchitis virus, laying hens, isolates, phylogeny, vaccines.
Introducción
La industria avícola es el sector productivo de mayor crecimiento en el mundo, contribuyendo con la nutrición humana y generando ingresos para las economías familiares (Farrell, 2013). En Ecuador tiene gran proyección económica, debido a que la carne de pollo y los huevos de gallinas son fuentes de nutrientes con precios accesibles en el mercado (Aillón, 2012; Corporación Nacional de Avicultores del Ecuador -CONAVE-, 2020a). El consumo de huevos per cápita en el año 2018 fue de 218 unidades y ascendió a 226 por persona/año durante 2019 (Corporación Nacional de Avicultores del Ecuador -CONAVE-, 2020b). Según el censo avícola del año 2015, existen 310 granjas de aves ponedoras comerciales que se ubican mayoritariamente en la región del centro (Pastaza, Cotopaxi, Tungurahua y Chimborazo) y en la provincia de Manabí (Instituto Nacional de Estadística y Censos -INEC-, 2015). No obstante, la provincia de Tungurahua es la que cubre el 45% de la demanda de huevos de mesa en el país (Ministerio de Agricultura y Ganadería -MAG-, 2021).
El crecimiento de la avicultura ecuatoriana es resultado del trabajo colaborativo de los diferentes eslabones de esta cadena productiva (Agencia Ecuatoriana de Aseguramiento de la Calidad del Agro -AGROCALIDAD-, 2013). Sin embargo, el progreso de la avicultura puede verse afectado por enfermedades que influyen negativamente en la producción y, en consecuencia, la identificación y el control de las mismas resultan fundamentales para evitar pérdidas económicas (Bermudez, 2008).
La bronquitis infecciosa aviar es una de las enfermedades respiratorias más frecuentes y problemáticas dentro de las granjas de aves ponedoras comerciales en el mundo (Alexander & Senne, 2008; Cavanagh & Gelb, 2008). El agente causal es el IBV (Infectious bronchitis virus) perteneciente al género Gammacoronavirus de la subfamilia Orthocoronavirinae, familia Coronaviridae y orden Nidovirales (International Committee on Taxonomy of Viruses -ICTV-, 2020). El IBV es pleomórfico y posee un genoma de ARN de cadena simple con polaridad positiva y de longitud aproximada de 27,6 kilobases (Feng et al., 2017; Rafique et al., 2018). El genoma codifica para cuatro proteínas estructurales: la proteína de espícula (S), la glicoproteína de membrana (M), la proteína de la nucleocápside (N) y la pequeña proteína de membrana (E) (Abro, 2013; Marandino, 2013). Además incluye, en sus extremos, regiones no traducidas 5’ y 3’ UTR de baja variación, por lo que se suelen utilizar para determinar la presencia del IBV en muestras de granjas (Acevedo-Beiras, 2017). La entrada del IBV a la célula hospedadora se produce debido a la unión de la proteína S de la envoltura viral a los receptores específicos de la membrana celular (Shang et al., 2018). Esta proteína S se sintetiza como un precursor proteico (S0) de 1162 aminoácidos, que por modificación postraduccional genera dos subunidades funcionales, S1 y S2 (Abro, 2013; Liu et al., 2009a; Rafique et al., 2018). Las regiones hipervariables de S1 poseen una alta tasa de mutación, siendo no solo responsables de la antigenicidad viral, sino también de la mayoría de las variaciones genéticas del virus (Acevedo-Beiras, 2010; Dolz et al., 2008; Montassier, 2010).
Debido a la existencia de diversas cepas del IBV, la instauración de un programa vacunal efectivo para la prevención y control de la bronquitis infecciosa constituye un verdadero desafío (de Wit et al., 2011). La información científica en Ecuador se limita a escasas publicaciones enfocadas al monitoreo en las aves de traspatio mediante la utilización de las técnicas de ELISA en el noroeste, hemoaglutinación en Galápagos y PCR en tiempo real en la parroquia de Puéllaro (García, 2016; Hernandez et al., 2006; Soos et al., 2008). Las investigaciones mencionadas y las vacunas empleadas en Ecuador demuestran la presencia del IBV; sin embargo, no existen publicaciones científicas que expongan de manera fehaciente la presencia de las variantes que pueden estar circulando. Por lo tanto, el objetivo de la investigación fue caracterizar, mediante técnicas moleculares, las cepas del IBV circulantes en las explotaciones de gallinas ponedoras de la provincia de Tungurahua.
Materiales y métodos
Muestreo
Sobre la base de los datos del censo avícola disponible, se incluyeron, al azar, 47 granjas. Para realizar un muestreo homogéneo se generaron clústeres con la georreferenciación de las granjas, tomando en cuenta zonas de tolerancia de 2 km. La creación de clústeres se realizó con el software QGIS 3.18.1 (Figura 1). El muestreo se realizó entre noviembre de 2020 y agosto de 2021.
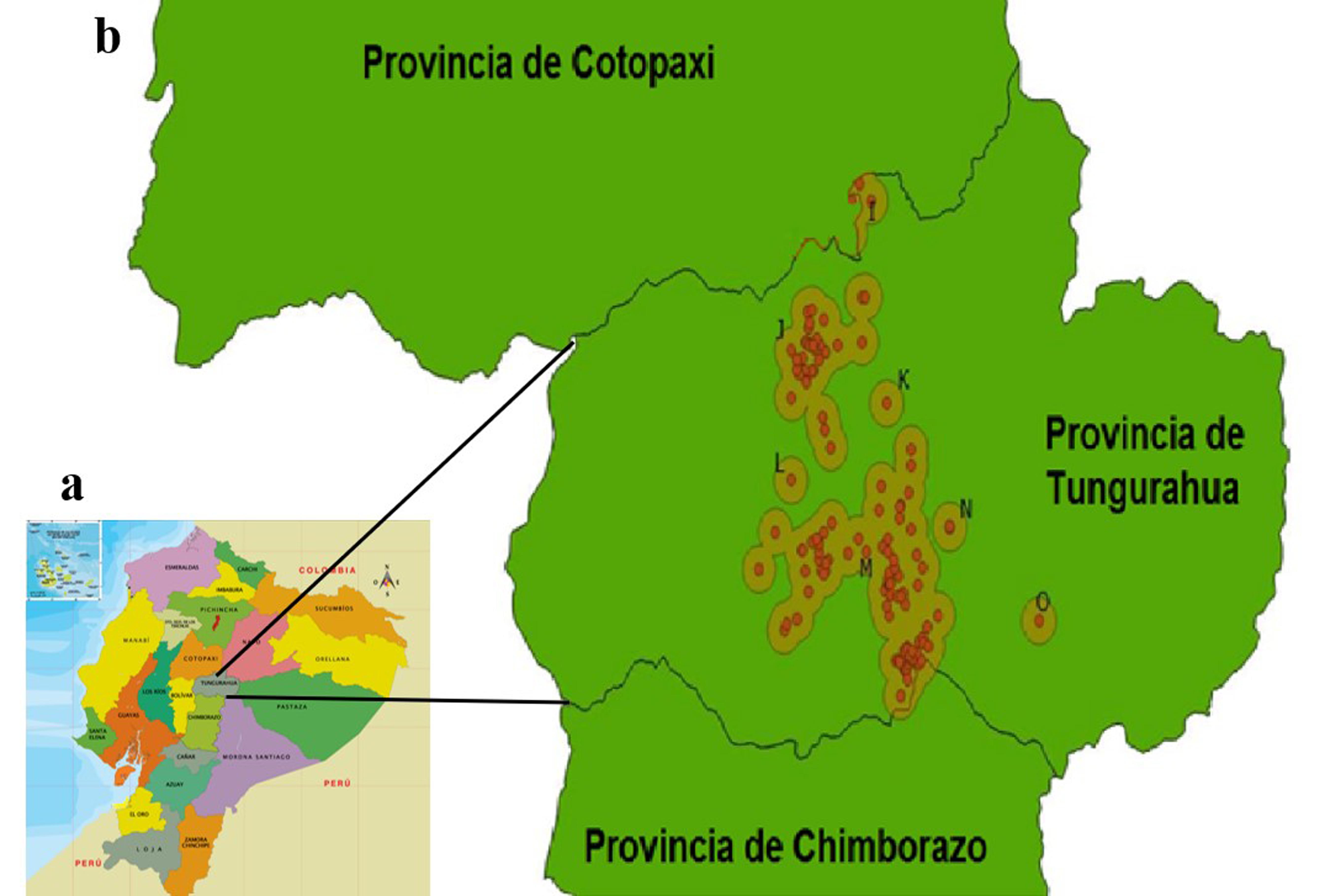
Clústeres para muestreo del virus de la bronquitis infecciosa aviar (IBV) en la provincia de Tungurahua. a. Mapa político del Ecuador (https://www.prolipa.com.ec/actividades/mapapolitico). b. Imagen generada en QGIS 3.28.1 a partir de las coordenadas geográficas de las granjas avícolas de la provincia de Tungurahua que se identifican con círculos. Se observan siete clústeres identificados con letras de la I a la O.
De cada granja se seleccionó un galpón con aves de 15 a 100 semanas de edad y de cada galpón se tomaron cinco aves al azar, las que fueron transportadas hasta la Facultad de Medicina Veterinaria y Zootecnia de la Universidad Central del Ecuador.
Se tomaron hisopados cloacales y laringotraqueales de todas las aves y luego se sacrificaron mediante electrocución de una sola aplicación (FCV-UNLP-CICUAL 124-5-22T). Se tomaron, además, muestras de tráquea, pulmón, oviducto y tonsilas cecales, las que fueron colocadas en frascos individuales con solución tamponada de fosfato estéril con antibióticos (penicilina 10.000 U/ml, estreptomicina 10.000 µg/ml, Eurobio SA) y posteriormente almacenadas a -80 °C. Se escogieron, además, 17 vacunas a virus vivo, usadas en la zona de estudio, para comparar sus secuencias con aquellas obtenidas a partir de las muestras de campo.
Procesamiento de las muestras
Las muestras de hisopados se descongelaron de manera rápida a 37 °C en baño María, se centrifugaron a 4500 rpm durante 30 minutos a 4 °C y se tomaron 1,5 ml de cada uno de los sobrenadantes para ser conservados a -80 °C. Las muestras de órganos se descongelaron como fue descripto, se cortaron en trozos pequeños y se molieron y homogeneizaron con medio mínimo esencial estéril (MEM MP Biomedicals) para lograr suspensiones al 20% p/v. Estas suspensiones se centrifugaron a 4500 rpm durante 30 minutos a 4 °C y 0,5 ml de cada uno de los sobrenadantes obtenidos se agruparon de acuerdo al siguiente criterio: a) muestras de aparato respiratorio (hisopado laringotraqueal, suspensión de tráquea y pulmón); b) muestras de aparato reproductivo y sistema inmune (suspensión de oviducto y tonsilas cecales) y c) hisopado cloacal.
Se realizó, además, la reconstitución de las vacunas mencionadas en la Tabla 1, con el diluyente correspondiente y según las indicaciones del fabricante.
| No | Código para vacuna | Cepa del IBV | |
| 1 | 1B | Ma 5 | |
| 2 | 2B | 4/91 | |
| 3 | 3B | B48 | |
| 4 | 4B | Variantea | |
| 5 | 6B | H120 | |
| 6 | 7B | H120 | |
| 7 | 8B | Massa | |
| 8 | 9B | H120 | |
| 9 | 10B | Massa | |
| 10 | 1NB | H120 | |
| 11 | 2NB | Massa | |
| 12 | 3NB | Massa | |
| 13 | 4NB | Massa | |
| 14 | 5NB | Tipo Massa | |
| 15 | 10NB | Tipo Mass a | |
| 16 | 11NB | Ma5 | |
| 17 | 14NB | Mass |
Referencias: a La descripción del fabricante solo menciona cepa Mass, tipo Mass o variante
Técnicas moleculares
Extracción de ARN y transcripción reversa
El ARN de cada grupo de muestras (a, b y c), así como el de las vacunas, fue extraído por separado mediante el uso del kit comercial QIAmp® Viral RNA Mini Kit (QIAGEN). Para la extracción se utilizaron 140 µl del líquido sobrenadante y se siguieron los pasos mencionados en el instructivo del fabricante.
A partir de los 80 µl de ARN obtenido se procedió con la retrotranscripción a ADNc utilizando para ello el OneScript Plus cADN Synthesis Kit (abm, G236). Se siguió el protocolo establecido en las indicaciones, empleando 25 ng/µl como concentración final mínima y oligonucleótidos al azar.
Se realizó un control interno de extracción y retrotranscripción mediante PCR para una región del exón 3 de la β-actina aviar. Se utilizaron los cebadores FW 685 (5’- GAGAAATTGTGCGTGACATCA-3’) y RV 818 (5’-CCTGAACCTCTCATTGCCA- 3’) para amplificar un segmento de 152 pb (Marandino, 2013). La prueba para β-actina se llevó a cabo con cada tipo de muestra (a, b y c).
Detección del IBV mediante PCR para la región 5’UTR
Para amplificar un segmento de 143 pb de la región 5’UTR del IBV se utilizaron los siguientes cebadores: IBV5’ GU391 (5’-GCTTTTGAGCCTAGCGTT-3’) y IBV5’ GL533 (5’-GCCATGTTGTCACTGTCTATTG-3’) (Callison et al., 2006).
Para realizar los ensayos de PCR se utilizó la enzima polimerasa GoTaq® Flexi DNA Polymerase (PROMEGA, M9295). Se trabajó inicialmente con las condiciones de termociclado recomendadas por el fabricante de la polimerasa, en un volumen total de reacción de 15 µl.
Como control positivo se usó ADNc de IBV facilitado por la Agencia de Regulación y control Fito y Zoosanitario del Ecuador (AGROCALIDAD).
También se procedió a detectar la presencia del IBV en cada una de las 17 vacunas a virus vivo.
PCR para un segmento del gen codificante de la proteína S1 del IBV
Se consideraron como positivas las granjas con, al menos, una muestra que amplificó el segmento de la región 5’UTR. Solo una muestra positiva a IBV de cada granja fue evaluada mediante PCR para la región S1. Del mismo modo, se amplificaron las muestras de vacunas, con cebadores previamente diseñados para caracterizar cepas del IBV en Argentina y Uruguay (Marandino, 2013). El par de cebadores utilizados fue: IBV-S1 5’-ACTGAACAAAAGACAGACTT-3’ e IBV-S2 5’-CCATCTGAAAAATTRCCAGT-3’, que amplifica un fragmento de 783 pb (Marandino, 2013; Marandino et al., 2015).
Para la reacción de PCR se utilizó una desnaturalización inicial a 94 °C durante 5 minutos, seguida de 40 ciclos de: desnaturalización a 94 °C durante 1 minuto, hibridación a 53 °C durante 1 minuto, extensión a 72 °C durante 1 minuto, y una extensión final a 72 °C durante 10 minutos. Se utilizó GoTaq® Flexi DNA Polymerase (PROMEGA M9295) a diferentes concentraciones (1,25U, 1,50U y 1,75U) y dNTP Mix (PROMEGA, U1515). Se ensayaron, además, diferentes concentraciones finales de cebadores (0,2 mM y 0,4 mM). El volumen final de reacción fue de 45 µl con 6 µl de muestra, 1,25 U de GoTaq y 0,2 mM de cebadores. Se verificó la especificidad de la técnica ensayando la PCR con dos vacunas a virus vivo contra la enfermedad de Newcastle (EN), ADNc del virus de la EN cedido por AGROCALIDAD y ADN del virus de Laringotraqueitis infecciosa aviar, también entregado por AGROCALIDAD. Las reacciones de PCR se realizaron en un termociclador SIMPLI AMPTM TERMAL CYCLER (APPLIED BIOSYSTEMS).
La electroforesis de los amplicones se realizó en una cuba horizontal ENDURO GelXL (LABNET), utilizando buffer TAE (40mM, EDTA 0,5M pH 8,0) a 120V y 400mA durante 60 minutos. El resultado de la migración electroforética en geles de agarosa (PROMEGA) al 2% se observó en un transiluminador UV (LABNET) y se registró con el uso de una cámara fotográfica digital.
Análisis filogenético
Los amplicones obtenidos para el fragmento del gen S1 del IBV fueron enviados para secuenciación a Macrogen Inc. (Corea). Las secuencias de nucleótidos (nt) fueron alineadas con los programas bioinformáticos MEGA-X 11.0.8. y Geneious Prime 2021.2.2. El alineamiento de las secuencias se realizó con el algoritmo de Clustal Omega. Para el análisis filogenético se emplearon secuencias parciales de la subunidad S1 (desde el codón de inicio ATG hasta el sitio 664 pb). Se incluyeron, además de las secuencias de las muestras y las 17 secuencias vacunales logradas durante la investigación, 32 cepas prototipos de la clasificación genotípica de Valastro et al. (2016) (Tabla 2) y 32 secuencias consideradas como representativas para América, Europa y China (Tabla 3).
El árbol filogenético se construyó empleando el programa IQ-Tree (https://www.hiv.lanl.gov/content/sequence/IQTREE/iqtree.html). IQ-Tree realizó automáticamente los cálculos y estimó el mejor modelo a través de ModelFinder (Kalyaanamoorthy et al., 2017; Nguyen et al., 2015). Además, el programa construyó un árbol consenso de máxima similitud con 1000 ultra-bootstrap para el conjunto de datos (Hoang et al., 2017).
| Código de acceso al GenBank | Cepa prototipo | Código en árboles filogenéticos | País o procedencia | Linaje |
| M95169 | BEAUDETTE | Beaudette_GI-1 | Estados Unidos | GI-1 |
| GU393336 | HOLTE | Holte_GI-2 | Estados Unidos | GI-2 |
| L14069 | GRAY | Gray_GI-3 | Estados Unidos | GI-3 |
| L18988 | HOLTE | Holte_GI-4 | Estados Unidos | GI-4 |
| U29522 | N1/62 | N1/62_GI-5 | Australia | GI-5 |
| U29519 | VicS | VicS_GI-6 | Australia | GI-6 |
| AY606320 | TP/64 | TP/64_GI-7 | Taiwán | GI-7 |
| JQ964061 | L165 | L165_GI-8 | Estados Unidos | GI-8 |
| M99482 | ARK99 | Ark99_GI-9 | Estados Unidos | GI-9 |
| AF151954 | B | B_GI-10 | Nueva Zelanda | GI-10 |
| JX182775 | UFMG/G | UFMG/G_GI-11 | Brasil | GI-11 |
| X52084 | D3896 | D3896_GI-12 | Países Bajos | GI-12 |
| EU914938 | Morroccan-G/83 | Morroccan-G/83_GI-13 | Marruecos | GI-13 |
| X87238 | B1648 | B1648_GI-14 | Bélgica | GI-14 |
| FJ807932 | B4 | B4_GI-15 | Corea | GI-15 |
| KJ941019 | IZO28/86 | IZO28/86_GI-16 | Italia | GI-16 |
| AF419315 | CA/MACHADO/88 | CA/Machado/88_GI-17 | Estados Unidos | GI-17 |
| AY296744 | JP8127 | JP8127_GI-18 | Japón | GI-18 |
| KC577395 | 58HEN-93II | 58HEN-93II_GI-19 | China | GI-19 |
| AF349621 | Qu_mv | Qu_mv_GI-20 | Canadá | GI-20 |
| DQ064806 | SPAIN/97/314 | España/97/314_GI-21 | España | GI-21 |
| KC577382 | 40GDGZ-971 | 40GDGZ-971_GI-22 | China | GI-22 |
| AF093796 | VARIANT 2 | Variant_2_GI-23 | Israel | GI-23 |
| KF757447 | V13 | V13_GI-24 | India | GI-24 |
| EU925393 | CA/1737/04 | CA/1737/04_GI-25 | Estados Unidos | GI-25 |
| FN182243 | NGA/B401/2006 | NGA/B401/2006_GI-26 | Nigeria | GI-26 |
| GU301925 | GA08 | GA08_GI-27 | Estados Unidos | GI-27 |
| M21971 | D1466 | D1466_GII-1 | Países Bajos | GII-1 |
| U29450 | N1/88 | N1/88_GIII-1 | Australia | GIII-1 |
| U77298 | DE/072/92 | DE/072/92_GIV-1 | Estados Unidos | GIV-1 |
| DQ059618 | N4/02 | N4/02_GV-1 | Australia | GV-1 |
| GQ265948 | TC07-2 | TC07-2_GVI-1 | China | GVI-1 |
| Código de acceso al GenBank | Código en árboles filogenéticos | País o procedencia |
| MG734788.1 | CK/CH/GX/NN17-13 | China |
| MG734786.1 | CK/CH/GX/NN17-12 | China |
| MG734775.1 | CK/CH/CQ/17-2 | China |
| MG734773.1 | CK/CH/SC/DY17-1 | China |
| KU736751.1 | H120 cepa vacunal | Estados Unidos |
| KU736747.1 | MA5 cepa vacunal | Estados Unidos |
| DQ064815.1 | España/00/339 | España |
| DQ064812.1 | España/99/326 | España |
| DQ064809.1 | España/99/316 | España |
| KM658255.1 | AR/11/BA/28 | Argentina |
| KM658253.1 | AR/10/BA/30 | Argentina |
| KM658252.1 | AR/09/BA/39 | Argentina |
| KM658251.1 | AR/09/BA/38 | Argentina |
| EU283081.1 | Mass cepa vacunal | Estados Unidos |
| EU283060.1 | Conn cepa vacunal | Estados Unidos |
| EU283053.1 | Ark cepa vacunal | Estados Unidos |
| KM658245.1 | UY/09/CA/01 | Uruguay |
| KM658243.1 | UY/12/CA/39 | Uruguay |
| KM658241.1 | UY/12/CA/33 | Uruguay |
| KM658239.1 | UY/12/CA/27 | Uruguay |
| KY465751.1 | IBV/Brasil/ROL/0115 | Brasil |
| KY465748.1 | IBV/Brasil/NUP/0516 | Brasil |
| KY565555.1 | IBV/Brasil/FRQ/0315 | Brasil |
| KY565554.1 | IBV/Brasil/FRQ/0116 | Brasil |
| AF286302.1 | Q1/China | China |
| AF093794.1 | 4/91 cepa vacunal | Estados Unidos |
| Z83979.1 | UK/7/93 | Reino Unido |
| HM132098.1 | QX_UK | Reino Unido |
| MF043938.1 | 12.124_Chile | Chile |
| MF043935.1 | Q1_Chile | Chile |
| MF043933.1 | D100-273_Chile | Chile |
| GQ253482.1 | QX_España | España |
Resultados
El control interno de extracción y retrotranscripción funcionó correctamente y todas las muestras evaluadas resultaron positivas para ß-actina aviar (Figura 2). Por otro lado, 16 granjas (34,04%) resultaron positivas a IBV al registrar, al menos, una de las tres muestras empleadas que amplificara para la región 5’UTR (Tabla 4) (Figura 2). Del mismo modo, las 16 granjas resultaron positivas a la amplificación del segmento S1 (Tabla 4) (Figura 3).
| Código de granja | Resultados por tipo de muestra | ||||||
| No | 5'UTR | S1 | |||||
| a | b | c | a | b | c | ||
| 1 | U5240 | + | + | + | NA | + | NA |
| 2 | U5241 | + | + | + | NA | + | NA |
| 3 | U5247 | - | + | + | NA | + | NA |
| 4 | U5250 | + | - | - | + | NA | NA |
| 5 | U5256 | - | + | - | NA | + | NA |
| 6 | U5261 | - | + | - | NA | + | NA |
| 7 | U5262 | - | + | - | NA | + | NA |
| 8 | U5263 | - | + | - | NA | + | NA |
| 9 | U5264 | - | + | + | NA | + | NA |
| 10 | U5268 | + | + | + | NA | + | NA |
| 11 | U5270 | - | - | + | NA | NA | + |
| 12 | U5271 | + | + | + | NA | + | NA |
| 13 | U5272 | + | - | - | + | NA | NA |
| 14 | U5280 | - | + | - | NA | + | NA |
| 15 | U5281 | - | + | - | NA | + | NA |
| 16 | U5284 | - | + | + | NA | + | NA |
| Total | 6 | 13 | 8 | 2 | 13 | 1 | |
Referencias: a: hisopado laringotraqueal y suspensión de tráquea y pulmón; b: suspensión de oviducto y tonsilas cecales; c: hisopado cloacal.
+: Positivo, -: Negativo, NA: No analizado
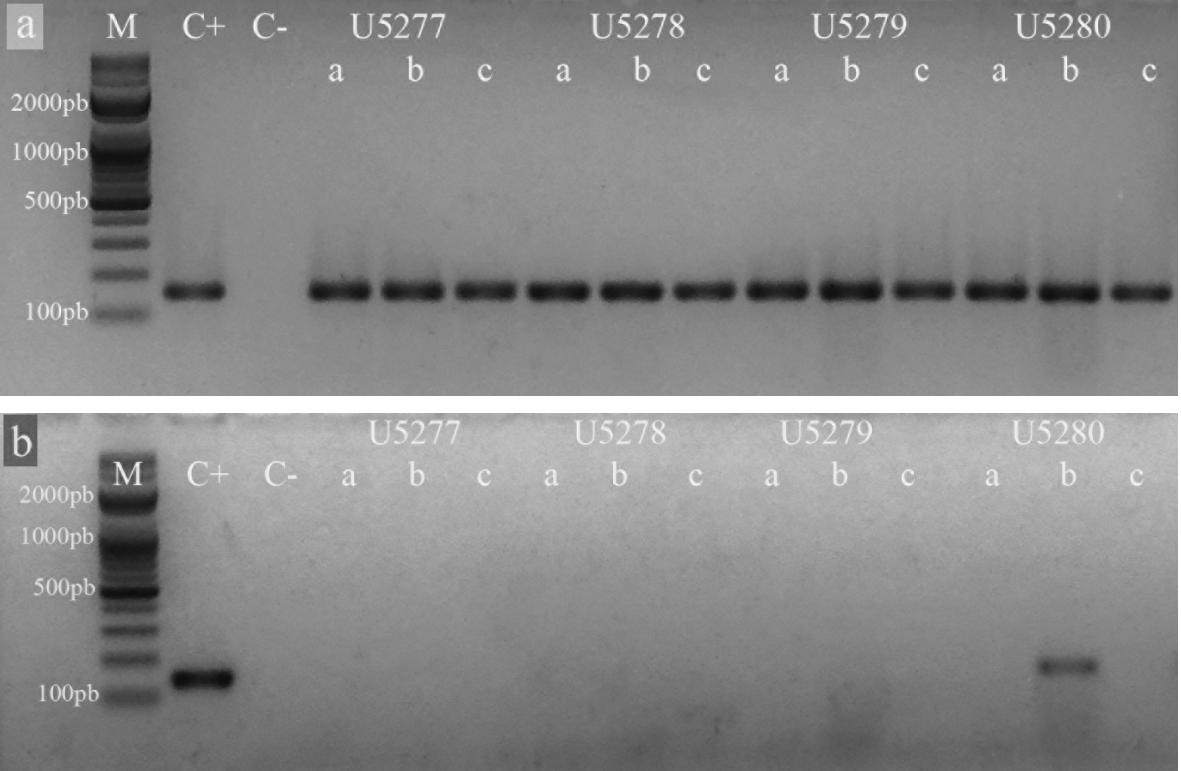
Técnica de RT PCR realizadas con muestras de cuatro granjas de la provincia de Tungurahua, Ecuador. Electroforesis en gel de agarosa al 2% teñidas con SYBR Safe DNA gel stain (INVITROGEN). a. amplificación de un segmento de 152 pb de la β-actina aviar. b. amplificación de un segmento de 143 pb de la región 5’UTR del virus de la bronquitis infecciosa aviar (IBV). Referencias: M: marcador de pares de bases, C+ control positivo, C- control negativo.
Con el servidor en línea gratuito IQ-TREE el mejor modelo que se obtuvo para el conjunto de secuencias fue: TPM3u+F+I+G4. A partir de este árbol consenso generado con IQ-TREE (Figura 4) se pudo determinar que cinco (U5250, U5261, U5272, U5281, U5284) de las 16 secuencias obtenidas a partir de las muestras se ubicaron en el mismo clado donde estaban presentes las cepas de tipo Massachusetts del linaje GI-1 (Figura 4). Por otro lado, siete de las secuencias recuperadas (U5240, U5241, U5263, U5264, U5268, U5271, U5280) se agruparon filogenéticamente en el clado del linaje GI-13 de las cepas tipo 793B. Finalmente, cuatro secuencias (U5247, U5256, U5262, U5270) de las granjas estudiadas se agruparon en un clado donde se ubicaron tanto cepas asiáticas como latinoamericanas del tipo Q1 del linaje G1-16 (Figura 4). Con relación a las vacunas comerciales analizadas, se observó que 15 estaban agrupadas en el linaje GI-1, en el que se ubican las cepas tipo Massachusetts, y solo dos vacunas (2B y 4B) se situaron en el mismo clado de las cepas vacunales de tipo 793B (Figura 4).
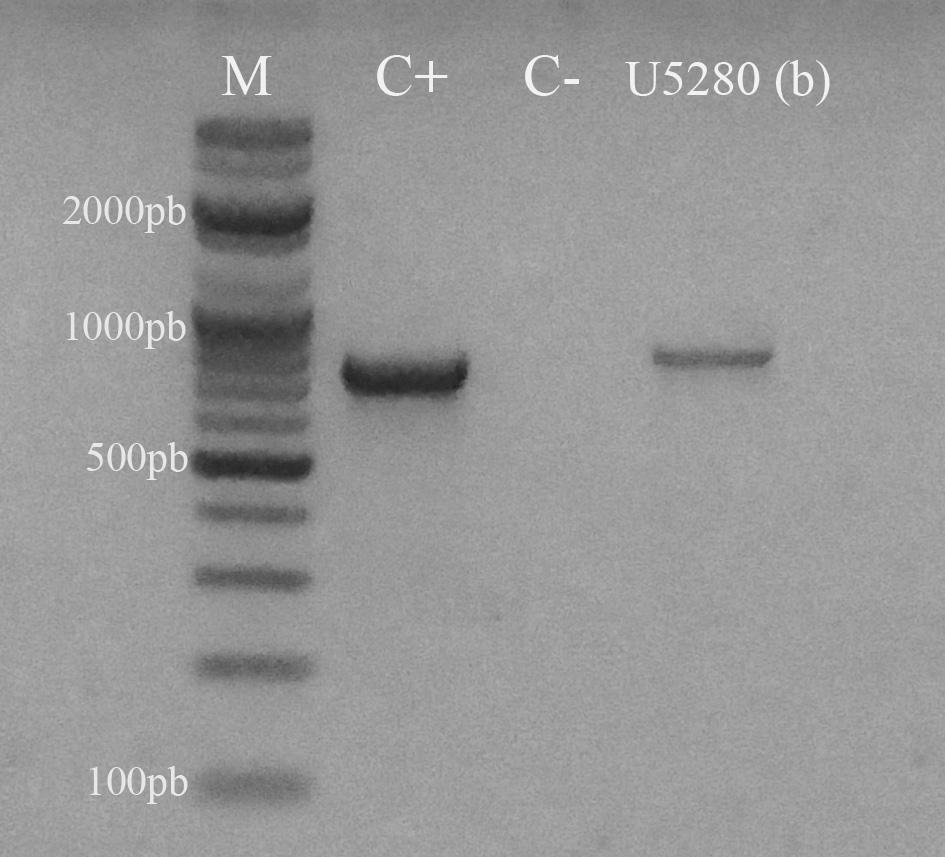
Gel de agarosa al 2% teñido con SYBR Safe DNA gel stain (INVITROGEN). Se observa la amplificación de un segmento de 783 pb del gen codificante de la proteína S1 del virus de la bronquitis infecciosa aviar (IBV). La muestra analizada correspondió a la codificada como U5280 y que resultó positiva a la región 5’UTR. Referencia. M: marcador de pares de bases, C+ control positivo, C- control negativo
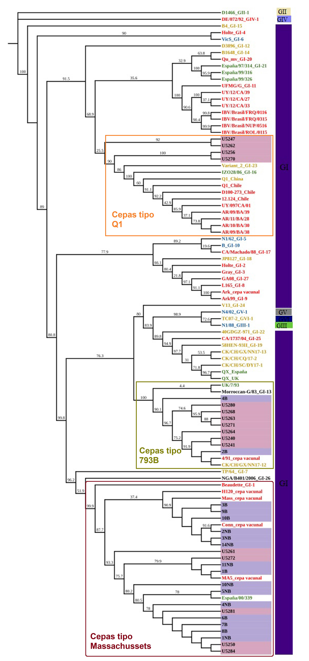
Árbol filogenético consenso con modelo TPM3u+F+I+G4. Los colores de letras hacen referencia a la procedencia de las cepas: rojo (América), amarillo (Asia), verde (Europa), celeste (Oceanía) y negro (África). Las cepas muestrales se observan en color negro y destacadas en recuadros rosa y las cepas de vacunas comerciales utilizadas en la zona de muestreo se muestran en color negro y ubicadas en recuadros lila. Los números a lo largo de las ramas hacen referencia a los valores de ultra-bootstrap.
Discusión y conclusiones
El análisis filogenético realizado mostró que cinco de las cepas encontradas en la provincia de Tungurahua correspondían a cepas vacunales de tipo Massachusetts. Es importante mencionar que las cepas vacunales de tipo Massachusetts son las recomendadas para utilizar en Ecuador (Organización Mundial de Sanidad Animal -OIE-, 2018). En un análisis filogenético realizado con aislamientos obtenidos en Uruguay entre 2009 y 2012 y en Argentina entre 2009 y 2011 se concluyó que las cepas no agrupaban con la cepa vacunal Massachusetts, la que también era la única autorizada para vacunación en estos países durante el desarrollo del trabajo (Marandino, 2013). Se debe mencionar que las muestras con las que trabajó esta investigadora provenían de brotes de infección por IBV y las muestras del presente estudio procedieron de aves que no manifestaron signología relacionada con IBV, pero que, al igual que en Argentina y Uruguay, habían sido vacunadas con la cepa Massachussets. Por su parte, Callison et al. (2006), también usando muestras de brotes de bronquitis infecciosa para amplificar la subunidad S1, concluyeron que los aislados estaban altamente relacionados con la cepa Massachusetts y que existía la probabilidad de detección y aislamiento de cepas vacunales. Del mismo modo, estudios de epizootiología molecular realizados en Suecia revelaron una fuerte probabilidad de que la vacunación con la cepa Massachussets había llevado a la propagación del virus vacunal, inclusive hacia una zona libre de vacunación (Farsang et al., 2002). En consecuencia, resulta razonable considerar que el virus circulante identificado en la provincia de Tungurahua sea el virus vacunal o cepas homólogas.
Siete de las secuencias obtenidas se encontraron en el clado del linaje GI-13 (las cepas tipo 793B) (Valastro et al., 2016). Según Valastro et al. (2016) la cepa Morroccan-G/83, aislada en Marruecos en 1983, sería la cepa progenitora del linaje, a pesar de que la primera cepa conocida fue aislada en Europa. Actualmente, las cepas de este linaje se encuentran presentes en muchas partes del mundo (Picault et al., 1995). En particular en América del Sur, las cepas de este tipo fueron introducidas como vacunas con la cepa 4/91 (Eldemery et al., 2017). Las cepas analizadas en el presente estudio podrían tener un origen vacunal, dado que dos de las vacunas analizadas (2B y 4B) se encuentran en este clado. Aunque la OIE (2018) sugiere para América del Sur la utilización de la cepa vacunal Massachusetts, en Ecuador están registradas para uso otras cepas vacunales como Arkansas, 4/91 y QX (Agencia Ecuatoriana de Aseguramiento de la Calidad del Agro -AGROCALIDAD-, 2021). De esta manera, se podría explicar el origen de las cepas variantes tipo 793 o 4/91 encontradas en el estudio desarrollado, tomando en cuenta que las cepas emergentes podrían ser el producto de la recombinación entre cepas de campo y cepas utilizadas para la vacunación (Quinteros et al., 2016).
Por otro lado, se estableció que cuatro de las muestras obtenidas en la provincia de Tungurahua estaban asociadas con cepas tipo Q1. Estas cepas tipo Q1 fueron aisladas inicialmente en Asia (Liu et al., 2009b; Yu et al., 2001) y luego se pudo evidenciar su presencia en América del Sur (de Wit et al., 2017; Marandino et al., 2015). Por ejemplo, en el análisis filogenético basado en la región S1 y realizado a partir de aislados chilenos de los años 2008 y 2009 se encontraron cepas de genotipo Q1 (de Wit et al., 2017). En consecuencia, la introducción de la cepa asiática podría haber tomado una ruta indirecta desde mercados latinoamericanos, dado que Ecuador no tiene comercio de importación directo con Asia. Además, debe recordarse que nuevas cepas de IBV pueden surgir también como producto del proceso replicativo característico de los coronavirus (Jackwood, 2012; de Wit et al., 2011).
Este trabajo constituye el primer estudio de caracterización molecular y análisis filogenético realizado con cepas encontradas en gallinas de postura comercial de Ecuador. Se concluyó que hay tres tipos de cepas circulando en la provincia de Tungurahua, dos de tipo vacunal o sus homólogas y una de origen desconocido. Se requieren estudios moleculares en los que se incluya la amplificación del gen S completo, a fin de que, con base en los estudios filogenéticos actuales que incluyen toda esta proteína, se puedan obtener datos más concluyentes.
Agradecimientos
El presente trabajo se financió con fondos otorgados como parte del programa de becas doctorales de la Universidad Central del Ecuador. Agradecemos a los avicultores de la provincia del Tungurahua, al Instituto Ecuatoriano del Huevo (INEH), a los médicos veterinarios Juan Valle y Richard Tuapanta, a los técnicos José Luis Medina y Sofía De Janón de la Unidad de Investigación de Enfermedades Transmitidas por Alimentos y Resistencia a los Antimicrobianos (UNIETAR) de Facultad de Medicina Veterinaria y Zootecnia de la Universidad Central Ecuador. También se agradece al personal del Laboratorio de Virología (LAVIR) de la Facultad de Ciencias Veterinarias de la Universidad Nacional de La Plata.
El aporte de todos ellos fue de gran utilidad para llevar a cabo el estudio.
Declaración de conflicto de intereses
Todos los autores declaran que no existe conflicto de intereses, incluyendo las relaciones financieras, personales o de otro tipo que pudieran influir en el trabajo.
Bibliografía
Abro S. 2013. Molecular Characterization and Detection of Infectious Bronchitis Virus. Doctoral Thesis, Faculty of Veterinary Medicine and Animal Science, Swedish University of Agricultural Sciences. http://pub.epsilon.slu.se/9299/1/abro_s_121210.pdf
Acevedo-Beiras A. 2010. Bronquitis infecciosa aviar: diagnóstico y control. REDVET (Revista Electrónica de Veterinaria).11(3):1-23. https://redalyc.org/pdf/636/63613123019.pdf
Acevedo-Beiras A. 2017. Virus de la bronquitis infecciosa: un desafío para la avicultura. Revista de Salud Animal. 39(3):1-12. http://scielo.sld.cu/pdf/rsa/v39n3/rsa07317.pdf
Agencia Ecuatoriana de Aseguramiento de la Calidad del Agro (AGROCALIDAD). 2013. Programa Nacional Sanitario Avícola. Dirección de Sanidad Animal: Programas Específicos. [En línea] Disponible en: https://oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Self-declarations/Annexes/ANEXO_3.pdf [Consultado 03-02-2019].
Agencia Ecuatoriana de Aseguramiento de la Calidad del Agro (AGROCALIDAD). 2021. Listado de cepas vacunales registradas ante la Agencia. [En línea] Disponible en: https://bing.com/ck/a?!&&p=40f9f82db4f7f61cc32c7209a24ac7d87d1847334c792b2e0c96cb5ac6c5a12cJmltdHM9MTY1NTEyMjQzNiZpZ3VpZD0xMmRlN2UzNS00MjZhLTQ1MTctOGUzZS1kZDBiZDBlMmE0ZjUmaW5zaWQ9NTEyOQ&ptn=3&fclid=4c018656-eb12-11ec-b00d-80843e13383e&u=a1aHR0cHM6Ly93d3cuYWdyb2NhbGlkYWQuZ29iLmVjL3dwLWNvbnRlbnQvdXBsb2Fkcy8yMDIxLzA3L0xpc3RhZG9fQ2VwYXNfUmVnaXN0cmFkYXNfQWdyb2NhbGlkYWQueGxzeA&ntb=1 [Consultado 12-04-2022].
Aillón M. 2012. Propuesta e implementación de un proyecto comunitario que se dedicará a la crianza, producción y comercialización avícola en la parroquia de Ascázubi. Tesis para la obtención del título de Ingeniera en Contabilidad y Auditoría. Carrera de Contabilidad y Auditoría, Universidad Central del Ecuador. http://dspace.uce.edu.ec/bistream/2500/1473/1/t-uce-003-272.pdf
Alexander D, Senne D. Newcastle Disease, Other Avian Paramyxoviruses, and Pneumovirus Infections. En: Saif, YM., Fadly, AM., Glisson, JR., McDougald, LR., Nolan LK. 2008. Diseases of poultry. 12th. ed. Iowa, State University Press. pp. 75-116.
Bermudez A. 2008. Introduction. En: Saif YM, Fadly AM., Glisson JR, McDougald LR, Nolan LK. 2008. Diseases of poultry. 12th. ed. Iowa, State University Press. pp. 3-46.
Callison SA, Hilt DA, Boynton TO, Sample BF, Robison R, Swayne DE, Jackwood MW. 2006. Development and evaluation of a real-time Taqman RT-PCR assay for the detection of infectious bronchitis virus from infected chickens. Journal of Virological Methods. 138(1-2):60-65. https://doi.org/0.1016/j.jviromet.2006.07.018
Cavanagh D, Gelb J. Infectious Bronchitis. En: Saif YM, Fadly AM, Glisson JR, McDougald LR, Nolan LK. 2008. Diseases of poultry. 12th. ed. Iowa, State University Press. pp. 117-135.
Corporación Nacional de Avicultores del Ecuador (CONAVE). 2020a. Cada año en Ecuador aumenta consumo de pollo y huevos. Maíz y Soya. [En línea] Disponible en: http://maizysoya.com/lector.php?id=20200548&tabla=articulos. [Consultado 05-06-2021].
Corporación Nacional de Avicultores del Ecuador (CONAVE). 2020b. Importancia de la producción avícola en el contexto nacional. Maíz y Soya. [En línea] Disponible en: http://maizysoya.com/lector.php?id=20200585. [Consultado 05-06-2021].
de Wit JJ, Cook JK, van der Heijden H. 2011. Infectious bronchitis virus variants: A review of the history, current situation, and control measures. Avian Pathology. 40(3):223-35. https://doi.org/10.1080/03079457.2011.566260
de Wit JJ, Dijkman R, Guerrero P, Calvo J, Gonzalez A, Hidalgo H. 2017. Variability in biological behaviour, pathogenicity, protectotype and induction of virus neutralizing antibodies by different vaccination programmes to infectious bronchitis virus genotype Q1 strains from Chile. Avian Pathology. 46(6):666-75. https://doi.org/10.1080/03079457.2017.1346782
Dolz R, Pujols J, Ordoñez G, Porta R, Majo N. 2008. Molecular epidemiology and evolution of avian infectious bronchitis virus in Spain over a fourteen-year period. Virology. 374(1):50-9. https://doi.org/10.1016/j.virol.2007.12.020
Eldemery F, Li Y, Yu Q, Van Santen VL, Toro H. 2017. Infectious bronchitis virus does not protect against ark-type challenge. Avian Diseases. 61(3):397-401. https://doi.org/10.1637/11632-032017-ResNoteR
Farrell D. 2013. Función de las aves de corral en la nutrición humana. [En línea] Disponible en: http://fao.org/3/a-al714s.pdf. [Consultado 12-03-2019].
Farsang A, Ros C, Renström LHM, Baule C, Soós T, Belák S. 2002. Molecular epizootiology of infectious bronchitis virus in Sweden indicating the involvement of a vaccine strain. Avian Pathology. 31(3):229-36. https://doi.org/10.1080/03079450220136530
Feng K, Wang F, Xue Y, Zhou Q, Chen F, Bi Y, Xie Q. 2017. Epidemiology and characterization of avian infectious bronchitis virus strains circulating in southern China during the period from 2013-2015. Scientific Reports. 7(1):1-11. https://doi.org/10.1038/s41598-017-06987-2
García W. 2016. Detección de bronquitis infecciosa aviar mediante diagnóstico molecular en aves de traspatio. Tesis para obtener el título de Médico Veterinario y Zootecnista, Facultad de Medicina Veterinaria y Zootecnia, Universidad Central del Ecuador. http://dspace.uce.edu.ec/bitstream/25000/11984/1/T-UCE-0014-033-2016.pdf
Hernandez S, Villegas P, Prieto F, Unda JC, Stedman N, Ritchie B, Carroll R, Hernandez-Divers SJ. 2006. A survey of selected avian pathogens of backyard poultry in northwestern Ecuador. Journal of Avian Medicine and Surgery. 20(3):147-58. https://doi.org/10.1647/2005-015R.1
Hoang DT, Chernomor O, von Haeseler A, Quang Minh B, Sy Vinh L. 2017. UFBoot2: Improving the ultrafast bootstrap approximation. Molecular Biology Evolution. 35(2):518-22. https://doi.org/10.1093/molbev/msx281
Instituto Nacional de Estadística y Censos (INEC). 2015. Aves planteles avícolas_nac2015. Encuesta de Superficie y Producción Agropecuaria Continua (ESPAC) 2015. [En línea]. Disponible en: https://anda.inec.gob.ec/anda/index.php/catalog/750/download/14283. [Consultado 02-03-2018].
International Committee on Taxonomy of Viruses (ICTV). Changes to virus taxonomy and the Statutes ratified by the International Committee on Taxonomy of Viruses (2020). Archives of Virology. 2020 ;165(11):2737-48. PMID: 32816125.
Jackwood M. 2012. Review of infectious bronchitis virus around the world. Avian Diseases Digest. 56(4):634-41. https://doi.org/10.1637/10360-1022712-digest.1
Kalyaanamoorthy S, Minh BQ, Wong TKF, von Haeseler A, Jermiin LS. 2017. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nature Methods. 14(6):587-9. https://doi.org/10.1038/nmeth.4285
Liu S, Zhang X, Wang Y, Li C, Han Z, Shao Y, Li H, Kong X. 2009a. Molecular characterization and pathogenicity of infectious bronchitis coronaviruses: Complicated evolution and epidemiology in China caused by cocirculation of multiple types of infectious bronchitis coronaviruses. Intervirology. 52(4):223-34. https://doi.org/10.1159/000227134
Liu S, Zhang X, Wang Y, Li C, Liu Q, Han Z, Zhang Q, Kong X, Tong G. 2009b. Evaluation of the protection conferred by commercial vaccines and attenuated heterologous isolates in China against the CK/CH/LDL/97I strain of infectious bronchitis coronavirus. Veterinary Journal. 179(1):130-6. https://doi.org/10.1016/j.tvjl.2007.08.018
Marandino A. 2013. Diagnóstico y caracterización genética del virus de la bronquitis infecciosa aviar en la industria avícola regional. Tesis de Maestría PEDECIBA Biología. Sub área genética, Universidad de la República del Uruguay. https://colibri.udelar.edu.uy/jspui/bitstream/20.500.12008/4029/1/uy24-16716.pdf
Marandino A, Pereda A, Tomas G, Hernandez M, Iraola G, Craig MI, Hernandez D, Banda A, Villegas P, Panzera Y, Perez R. 2015. Phylodynamic analysis of avian infectious bronchitis virus in South America. Journal of General Virology. 96(6):1340-6. https://doi.org/10.1099/vir.0.000077
Ministerio de Agricultura y Ganadería (MAG). 2021. Ecuador celebra el día nacional del huevo. [En línea] Disponible en: http://agricultura.gob.ec/ecuador-celebra-el-dia-nacional-del-huevo/
Montassier HJ. 2010. Molecular epidemiology and evolution of avian infectious bronchitis virus. Revista Brasileira de Ciencia Avícola. 12(2):87-96. https://doi.org/10.1590/S1516-635X2010000200003
Nguyen LT, Schmidt HA, von Haeseler A, Minh BQ. 2015. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Molecular Biology and Evolution. 32(1):268-74. https://doi.org/10.1093/molbev/msu300
Organización Mundial de Sanidad Animal (OIE). 2018. Avian infectious bronchitis. [En línea] Disponible en: https://oie.int/fileadmin/Home/eng/Health_standards/tahm/2.03.02_AIB.pdf. [Consultado 28-06-2020].
Picault J, Drouin P, Lamandé J, Allée C, Toux J, le Coq H, Guittet M, Bennejean G. 1995. L’epizootie recente de bronchite infectieuse aviaire en France. Proceedings of the 1eres Journée de la Recherche Avicole. 177-9. https://journees-de-la-recherche.org/JRA/Contenu/Archives/1_JRA/53epizootieJRA1.pdf
Quinteros J A, Lee SW, Markham PF, Noormohammadi AH, Hartley CA, Legione AR, Coppo MJC, Vaz PK, Browning GF. 2016. Full genome analysis of Australian infectious bronchitis viruses suggests frequent recombination events between vaccine strains and multiple phylogenetically distant avian coronaviruses of unknown origin. Veterinary Microbiology. 197:27-38. https://doi.org/10.1016/j.vetmic.2016.11.003
Rafique S, Siddique N, Abbas MA, Shah AA, Sharif A, Naeem K. 2018. Isolation and molecular characterization of infectious bronchitis virus (IBV) variants circulating in commercial poultry in Pakistan. Pakistan Veterinary Journal. 38(4):365-70. https://doi.org/10.29261/pakvetj/2018.071
Shang J, Zheng Y, Yang Y, Liu C, Geng Q, Luo C, Zhang W, Li F. 2018. Cryo-EM structure of infectious bronchitis coronavirus spike protein reveals structural and functional evolution of coronavirus spike proteins. PLoS Pathogens. 14(4):1-19. https://doi.org/10.1371/journal.ppat.1007009
Soos C, Padilla L, Iglesias A, Gottdenker N, Bedon M, Rios A, Parker P. 2008. Comparison of pathogens in broiler and backyard chickens on the Galápagos Islands: Implications for transmission to wildlife. The Auk. 125(2):445-55. https://doi.org/10.1525/auk.2008.06235.
Valastro V, Holmes EC, Britton P, Fusaro A, Jackwood MW, Cattoli G, Monne I. 2016. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infection, Genetics and Evolution. 39:349-64. https://doi.org/10.1016/j.meegid.2016.02.015.
Yu L, Jiang Y, Low S, Wang Z, Nam J, Liu W, Kwang J. 2001. Characterization of three infectious bronchitis virus isolates from China associated with proventriculus in vaccinated chickens. Avian Diseases. 45(2):416-24. https://doi.org/10.2307/1592981.
