

Chemical composition and initial growth of soybean seedlings originated from seeds produced under flooding condition at R5 growth stage
Composição química e crescimento inicial de plântulas de soja oriundas de sementes produzidas em condição de alagamento no estádio de crescimento R5
Revista de la Facultad de Agronomía
Universidad Nacional de La Plata, Argentina
ISSN: 1669-9513
Periodicity: Semestral
vol. 121, no. 2, 2022
Received: 18 February 2022
Accepted: 21 April 2022
Corresponding author: angel.celente10@gmail.com
Abstract: This work aimed to evaluate the chemical composition and the initial growth of soybean seedlings originated from seeds produced under influence of soil flooding in the R5 growthstage. The experimental design was totally randomized, organized in factorial scheme being two soybean cultivars x three soil-flooding periods, and the traits were disposed in four repetitions. With flooding during growth stage R5, the thousand seeds was reduced independently of the cultivar. The shoot growth and length, shoot dry matter and root dry matter were negatively influenced by soil flooding at the R5 stage in both cultivars. Oxygen deficiency can cause different changes in the protein concentration in the roots and in the aerial part of soybean seedlings in the periods and between the cultivars. The concentration of amino acids was stimulated in roots and shoots of seedlings originated from seeds produced under hypoxic conditions when comparing the cultivars.
Keywords: Glycine max (L) Merr, stress, photosynthesis, seeds, hypoxia.
Resumo: Este trabalho teve como objetivo avaliar a composição bromatológica e o crescimento inicial de mudas de soja oriundas de sementes produzidas sob influência de alagamento do solo no estádio R5. O delineamento experimental foi inteiramente casualizado, organizado em esquema fatorial sendo duas cultivares de soja x três épocas de alagamento do solo, e os caracteres dispostos em quatro repetições. Com o alagamento no estádio R5, o número de mil sementes foi reduzido independentemente da cultivar. O crescimento e comprimento da parte aérea, matéria seca da parte aérea e matéria seca da raiz, foram influenciados negativamente pelo alagamento do solo no estádio R5 em ambas cultivares. A deficiência de oxigênio causo diferentes alterações na concentração de proteína nas raízes e na parte aérea das mudas de soja nos períodos e entre as cultivares. A concentração de aminoácidos foi estimulada em raízes e partes aéreas de mudas originadas de sementes produzidas em condições hipóxicas quando comparadas as cultivares.
Palavras-chave: Glycine max (L) Merr, estresse, fotossíntese, sementes, hipóxia.
INTRODUCTION
The soybean (Glycine max (L.) Merrill) is an important economic specie, due to being the leading source of vegetable oil and protein, and it is consolidated in Brazil, which is one of the largest worldwide producers (Santos et al., 2012; Szareski et al., 2018). It produces approximately 119 million tons of grains in an area that is superior to 35 million hectares at the 2018/2019 growing season (Conab, 2019). The productivity of this crop depends on its genetic potential and on the environmental factors that interfere in the crop development, above all, in the photosynthesis. The photosynthetic process depends on the radiation interception and on the conversion of it in chemical energy, being this dynamic influenced by many biotic and abiotic stresses, that can reduce the foliar area in sensitive growing and development stages (Lopes & Lima, 2015; Nardino et al., 2016; Gehlling et al., 2018; Rigo et al., 2018). Among them, stands out the soil draining unappropriated conditions that can compromise the crop productivity
The majority of world areas to the grains producing species presents soils with adequate draining conditions. However, approximately 6% of the terrestrial surface can suffer temporary flooding periods, particulary in Brazil there are about 33 million hectares of lowlands, which 5, 4 million hectares are in the Rio Grande do Sul state (Embrapa, 2005). Floods generat unfavorable environmental conditions for crop production, which intensifies with a greater frequency of rains, in this manner, is possible to happen a reduction on the oxygen availability in the roots, mainlydue to the slow oxygen diffusion on water (Zabalza et al., 2008). It can affect the growth of innumerous vegetal species (Coelho et al., 2013; Rosa et al., 2015). The oxygen deficiency due to flooding can compromise the physiologic aspects, the carbon assimilation, macronutrients absorption, as well, to suppress the roots respiratory metabolism (Dubal et al., 2016; Troyjack et al., 2018;).
Under flooding conditions, the oxygen availability to the cells reduces in order to maintain the aerobic respiration, which can result in energetic deficiency and compromise the vegetal metabolism (Bailey-Serres & Voesenek, 2008). The soil flooding severely interferes in the crops growth, being considered one of the main abiotic stresses. This cause a reduction in the seeds production and harm its chemical composition, physic and physiologic integrity (Wang et al., 2012; Szareski et al., 2016 & Follmann et al., 2016).
The alterations in the radicular system and the reduction in the photoassimilates translocation, as well by the reduction in the crops metabolic activity (Sachs & Vartapetian, 2007) due to soil flooding, cause the reduction on chlorophyll content and interfere in the efficiency and conversion of luminous radiation in chemical energy (Amarante et al., 2007).
The chemical composition in seeds embryos of several species has been studied by different authors. According to Ziegler (1995), the reserve proteins are used in the formation of structural compounds during the seedling growth. The seeds pre-formed reserves serve as respiration substrate during the pre-germination period and in the seedling development (Bewley & Black, 1994; Koch et al., 2018).
The soil flooding can occur in any phase of the vegetal development, but the negative effects are dependent of the genotype sensibility and the period of plants exposition to stress. Pedó et al. (2016), reports that, common bean seeds originated from plants that were submitted to temporary soil flooding compromise the germination and the vigor. Studies on the development of seeds produced under flooding conditions in the reproductive period are essential and can support in the vegetal performance description against the occurrence of environmental stress.
In this manner, the present work aimed to evaluate the chemical composition and the initial growth of soybean seedlings originated from seeds produced under influence of soil flooding in the crop reproductive period.
MATERIAL AND METHODS
Conduction of study
The experiment was carried out in a “PW” model greenhouse (chapel model) coated with polycarbonate, disposed in north-south direction and equipped with controlled temperature and air humidity, with geographic location at 31.52’S and 52.21’W, belonging to the Federal University of Pelotas in 2014/2015 harvest.
It soyabean seeds from the cultivars Fundacep 64 (C1) and Fundacep66 (C2) were used, disposed in black polyethylene containers, with 12 liters of capacity, containing a substrate composed with horizon A.soil, from a Solodic Planosol (Aridic Haplustalf), with fertility previously corrected according to the soil analysis and based on the Fertility and Liming Manual (CQFS RS/SC, 2004).
During stage R5 (correspondin gto the start of seed filling) the soil was temporarily flooded. In order to make possible the treatments establishment, the polyethylene containers, filled with soil, were perforated in the inferior part to facilitate the draining of water excess and to maintain the soil field capacity.
The field capacity was determined using the tension table methodology (Embrapa, 1997) and from it was defined the water volume necessary to establish the flooding by zero, two and four days In the flood treatment, a water blade of 20 mm was maintained on the soil surface by fitting a second black polyethylene container without perforations on top of the soil-filled containers. For the draining of the flooded soil, it was realized the remove of the container on the top, allowing the water draining until field capacity.
The C1 and C2 seeds harvest was manually realized with approximately 16% humidity. Later, it were dried with forced air kiln, with average temperature of 35ºC, stationary dryer, until homogeneity of 12% humidity, after 72 hours. The seeds were manually separated and stored in cold chamber with average temperature of 15ºC and air relative humidity of 60%.
Traits measured
For the evaluation of the influence of soil flooding in R., following evaluations were realized:
Thousand seeds weight was measured with eight samples of 100 seeds from the utile area of the experimental plot, randomly obtained and counted with manual counter and extrapolated to thousand seeds.
For the evaluation of seedlings growth, the seeds were submitted to proceedings adapted from AOSA (1983) and described by Nakagawa (1999) where, for the determination of shoot and root length of seedlings, were used four subsamples of 10 seedlings, after 21 days. The length of the shoot was obtained by the distance between the basal part of the upper part of the seedling and the length of the root was determined by the distance between the stem and the cap. Results were expressed in millimeters per seedling (mg seedling-1). The shoot and primary root dry matter were obtained from four subsamples of 10 seedlings. Therefore, the seedlings were conditioned in craft paper envelopes and submitted to dry in a forced air kiln, with 70ºC temperature until constant mass, approximately 72 hours. The results were expressed in milligrams per seedling (mg seedling-1).
The protein contents were quantified in samples of shoot and root fresh matter (MF) (using 100 mg of Comassie blue in 50 mL of ethanol (95%), being added 100 mL of H.PO. (85%) and completed for one liter. Afterwards, the material was constantly agitated and submitted to filtration (Bradford, 1976) and expressed in mg g-1 of MF.
For the seedlings amino acid content determination, the extracts were obtained according to Bieleski & Turner (1966) methodology with modifications. The samples with 0,250 mg of vegetal material were macerated with liquid nitrogen and homogenized with 10 ml of MCW extracting solution (methanol: chloroform: water; proportion of 12:5:3). After 24 hours, the extracts were centrifuged with 600 g per 30 minutes. For each 4 ml of supernatant 1,0 ml of chloroform and 1,5 ml of water, were added, and centrifuged for the phases separation. The supernatant containing the metabolites was collected and transferred for water-bath in 38ºC, remaining for 30 hours for the chloroform residue elimination and samples concentration for the total soluble amino acids analysis determined according to Yemm & Cocking (1955).
The seedling extracts were properly diluted in pure water, in a final volume of 1 ml in test tubes where 0,5 mL of citrate buffer 0,2 M with 5,0 pH and 0,2 ml of ninhydrin reactive 5% in methylcellusolve (monomethyl ether of ethylene glycol) and 1 ml of KCN 2% (v/v) in methylcellusolve (prepared from the solution 0,01 M of KCN in pure water) were added.
The test tubes were agitated, covered with glass balls and transferred to a water-bath on 100ºC for 20 minutes. As follows, the tubes were placed in the dark until achieving room temperature and then it was added 1,3 ml of ethanol 60%, where it was completed to 4 ml and realized the tubes agitation over again. The leucine optical density patterns (0-200 nmol) and the samples was measured at 570 nm, being realized three analytic repetitions for each sample and the content of total soluble amino acids was expressed in µmol g-1 MF.
Experimental design and statistical analysis
The experimental design was totally randomized, organized in factorial scheme being two soybean cultivars (Fundacep64:C1, and Fundacep66: C.) x three soil-flooding periods (0, 2 and 4 days), and they were disposed in four repetitions. The data was submitted to analysis of variance at 5% probability by F test, the significant interactions were separated in to simple effects, and the characters that did not revealed interaction were separated in to principal effects at 5% probability by Tukey test.
RESULTS
The variance analysis revealed significant interaction between cultivars x soil flooding periods at 5% probability for the characters thousand seeds weight (Table 1), shoot length, root length, shot dry matter (Table 2), and root protein (Table 3). The characters root dry matter, shoot amino acids, and shoot protein, did not presented significant interaction and were separated in to the principal effects thru complementary analysis.
The thousand seeds weight presented reduction due to the increment of soil flooding period (Table 1). For both cultivars, the higher thousand seeds weight (TSW) was obtained in the no-flooding period (0 days), however, for this period the higher TSWwas showed by cultivar 2 being superior in 54,96 grams.

The lowest TSW averages were obtained thru the flooding period of 4 days, with average of 133.66 g and 129.59 g for the C1 y C2, respectively. However, for the C1, treatments with 2 and 4 flooding day periods did not statistically differ between them but in the cultivar 2 did.
The shoot length originated from seed produced on flooded soil was negatively affected by the increment of the period of stress. For both cultivars, the shorter shoot length was obtained in 4 days of flooding with averages of 5.45 and 5.06 cm for the C1 y C2, respectively (Table 2). Still, the flooding periods 0 and 2 days did not statistically differ among themselves. For this variable, the higher averages in the periods 0 and 2 days of flooding were observed thru the C1, though, in the period of 4 days of flooding there was no significant difference between cultivars.
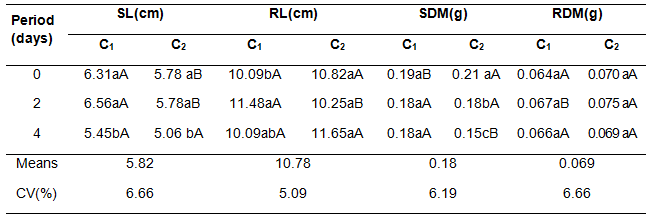
The average values of shoot dry matter of the seedlings coming from plants different flooding periods did not differ for the C1, however, the C2 presented higher average in periods with no-flooding (0 days), followed by the periods of 2 and 4 days, with average values of 0.219; 0.180 and 0.150 g, respectively (Table 2). For this variable the period with absence of flooding revealed higher average for the C2, while in the period of 4 days it was obtained increment in the C1. For root dry matter, at different times of flooding, no difference was observed in c1 and c2 respectively, but C2 in the period with 2 days of flooding showed an increase in root dry matter (Table 2).
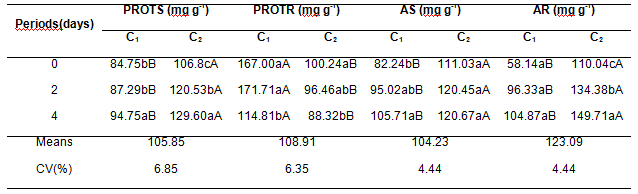
The protein content obtained in the shoots increased in plants submitted to higher periods of flooding in both cultivars. For the C1, the flooding period of 4 days presented higher protein concentration (94.75 mg g-1), meanwhile the lowest concentration was obtained in plants with no flooding soil. (Table3) The same behavior was observed for the C2, where there was an increment in the shoot protein content due to the in the longer flooding period, however, to relate the cultivar it was observed superiority to the C2, with increasing protein increment.
The protein contents accumulated in the roots of soybean seedlings presents distinct behavior when compared to the shoot. The highest contents of protein in the roots were evidenced in periods with lower flooding periods for both cultivars. However, the superiority for the C1 was observed in all flooding periods. There was, in this manner, alteration in the protein accumulation in the roots in relation to the leaves.
The highest amino acid content was observed for both structures in the C2 in all flooding periods (Table 3). The inundation of the radicular system did not affect the amino acid contents in the shoot of the C2 and in the roots of the C1. Conversely, there was an increment in the amino acid content in the roots of the C2 and in the shoot of the C1, where the highest amino acid content occurs in the period of 4 days of flooding.
DISCUSSION
The increase in the soil flooding period reduced the TSW, this seed physic attribute is related to the amount of assimilates allocated during its development, being that the seeds develop in more favorable environments and can result in higher mass of thousand seeds weight (Peske et al., 2012; Carvalho et al., 2016; Ferrari et al., 2018).
The synthesis and distribution of assimilates between plant structures is a preponderant factor for the production of high quality seeds. The occurrence of environmental stress can change the adequate distribution of dry matter between different plant structures. In this way, many environmental stresses have impacted CO2 assimilation, which can reduce electron transfer efficiency, negatively interfering with photosynthesis (Aumonde et al., 2017), and thus directly affecting the production of carbon compounds intended for plant growth and development (Batista et al., 2008). The reproductive period is characterized by its higher dry mass accumulation in the reproductive structures, where the abiotic stresses in this period can negatively affect the seeds filling, once there is a higher exigency of assimilates originated from the photosynthesis and of nutrients in the soybean reproductive period (Zimmer et al., 2016; Zanatta et al., 2018).
It is highlighted that the seeds mass depends on the size, duration and photosynthetic apparatus functioning, as well, the efficiency of the photo assimilates translocation, of the seeds filling period and the edaphoclimatic conditions during the development of the matrix plant and seed (Vieira Junior & Dourado Neto, 2008; Meira et al., 2016; Soares et al., 2016).
The higher dry matter of the seedlings structures coming from plants different flooding periods, can be related to the higher capacity of resources mobilization and assimilate allocation, that result in the seedling growth (Burgos et al., 2004; Monteiro et al., 2016; Troyjack et al., 2018). The flooding stress can cause reduction in the vegetal metabolism, lower reserve allocation, what can reduce its deposition in seeds and its quality (Gehlling et al., 2015), what afterwards, can affect the seedling performance.
The cultivars distinct behaviors, in front of the stress occurrence, might be related different tolerance of genotypes to soil flooding. This is linked to its capacity to develop biochemical, physiological and morphoanatomical answers, which can guaranty efficient mechanisms to maintain the growth, maintenance of vital functions to survive under hypoxia conditions (Aumonde et al., 2017).
Changes in the protein levels in the shoot of soybean seedlings indicate that the highest protein production is related to the tolerance to stress in hypox soil conditions (Shingaki-Wells et al., 2011; Perata et al., 2011; Pedó et al., 2016; Carvalho et al., 2017). Carbohydrate supply and regulation of carbohydrate and energy metabolism are important in overcoming hypoxic stress (Andreev et al., 1991; Kumutha et al., 2008) leading many plants to the accumulation of amino acids (Fan et al., 1988). Possibly, the increase of protein content in the plant due to the flooding reflects in the future generation of the seedlings, revealing higher amino acid concentration. Under flooding conditions the cellular O2 level can decrease to very low concentrations triggering a cascade of signalization/transduction of signals that alter the crops metabolism (Aumonde, 2017).
In general, the flooding in R. growth stage in soybean negatively affects physic characteristics of the species, as the thousand seeds. But also, it can influence the variables of initial growth and chemical composition in the seedlings of the next generation.
Finally, this study reports that, when submitting soybean plants to flooding at the R5 stage, the weight of a thousand seeds is reduced regardless of the cultivar. The shoot growth, length and dry matter were negatively influenced by soil flooding at the R5 growth stage in both cultivars. The dry matter of roots in the different times of flooding was not observed difference in the cultivars, only when purchased, there is evidence of an increase in the roots at 2 days in C2. Oxygen deficiency caused alterations in the protein concentration in the roots and in the aerial part of the soybean seedlings in a different way in the periods and between the cultivars. The amino acid content was stimulated in roots and shoots of seedlings originated from seeds produced under hypoxic conditions, showing differences between cultivars.
REFERENCES
Amarante, L., Colares, D.S., Oliveira, M.L., Zenzen, I.L., Badinelli, P.G., Bernardi, E. (2007). Chlorophyll content in soybean symbiotically associated with different strains of Bradyrhizobium under flooding. Revista Brasileira de Biociências, Porto Alegre, 5(2): 906-908.
Andreev, V.Y., Generozova, I.P., Vartapetian, B.B. (1991). Energy status and mitochondrial ultrastructure of excised pea root at anoxia and post-anoxia. Plant Physiology and Biochemistry, v.29, p. 171-176.
Association of Official Seed Analysts- AOSA (1983). Seed vigor testing and book. East Lansing: AOSA, 93 p.
Aumonde, T.Z., Pedó, T., Martinazzo, E.G., Villela, F.A. (2017). Estresses Ambientais na Produção De Sementes: Ciência e Aplicação. 1ª ed. Pelotas: Cópias Santa Cruz, v. 1. 313p.
Bailey-Serres J, Voesenek, L.A.C.J. (2008) Flooding stress: acclimations and genetic diversity. Annual Review of Plant Biology, Palo Alto, 59: 313-319.
Batista, C.U.N., Medri, M.E., Bianchini, E., Medri, C., Pimenta, J.A. (2008).Tolerância à inundação de Cecropia pachystachya Trec. (Cecropiaceae): aspectos ecofisiológicos e morfoanatômicos. Acta Botânica Brasileira, v.22, p.91-98.
Bewley, J.D., Black, M. (1994). Physiology of development and germination. New York: Plenum Press, 445p.
Bieleski, R.L., Turner, N. (1966). Separation and estimation of aminoacids in crude plant extracts by thin-layer eletrophoresis and chromatography. Analytical Biochemistry, 17: 278-293.
Bradford, M.M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72: 248-254.
Burgos, N.R., Talbert, R.E., Kim, K.S., Kuk, Y.I. (2004). Growth inibition and root ultrastructure of cucumber seedlings exposed to allele chemicals from rye. Journal of Chemical Ecology, 30(3): 671 - 690.
Carvalho, I.R., Nardino, M., Demari, G., Follmann, D.N., Regatti, A., Olivoto, T., Pelegrin, A.J., Szareski, V.J., Warths, C.A., Koch, F., Aisenberg, G.R., Pedó, T., Aumonde, T.Z., Souza, V.Q. (2016.) Multivariate analysis applied to genotype distinction in common bean. International Journal of Current Research, 8: 38352-38358.
Carvalho, I.R., Nardino, M., Demari, G.H., Szaresk, V.J., Follmann, D.N., Pelegrin, A.J., Ferrari, M., Olivoto, T., Barbosa, M.H., Oliveira, A.C., Maia, L.C., Souza, V.Q. (2017). Relations among phenotypic traits of soybean pods and growth habit. African Journal of Agricultural Research,12: 450-458.
Coelho, C.C.R., Neves, M.G., Oliveira, L.M., Conceição, A.G.C., Okumura, R.S., Oliveira, Neto. C.F. (2013). Biometry in maize plants submitted to flooding. Agroecossistemas, Marabá, 5(1): 32-38.
Companhia Nacional De Abastecimento (Conab) (2019). Acompanhamento de safra brasileira de grãos, safra 2018/2019. Brasília, 3(1).
Dubal, I., Troyjack, C., Koch, F., Aisenberg, G.R., Szareski, V.J., Pimentel, J.R., Nardin, M., Carvalho, I.R., Souza, V.Q., Villela, F.A., Aumonde, T.Z., Pedó, T. (2016). Effect of temperature on bean seed germination: vigor and isozyme expression. Agricultural Science Research Journals, 1: 001-009.
Embrapa. (1997). Manual de métodos de análise de solo. 2.ed. Centro Nacional de Pesquisa de Solos, Rio de Janeiro, 212p.
Embrapa (2005). Cultivo do Arroz Irrigado no Brasil. Embrapa Clima Temperado, Pelotas-RS. (Sistemas de Produção, 3).
Fan, T.W.M., Higashi, R.M., Lane, A.N. (1988). An in vivo 1H and 31P NMR investigation of the effect of nitrate on hypoxic metabolism in maize roots. Archives of Biochemistry and Biophysics, v. 266, p. 592-606.
Ferrari, M., Pelegrin, A.J., Carvalho, I.R., Nardino, M., Szareski, V.J., Olivoto, T., Rosa, T.C., Follmann, D.N., Pegoraro, C., Oliveira, A.C., Maia, L.C., Souza, V.Q. (2018). Path analysis and phenotypic correlation among yield components of soybean using environmental stratification methods. Aust J Crop Sci, 12: 193-202.
Follmann, DN., Souza, V.Q., Nardino, M., Carvalho, I.R., Schmidt, D., Ferrari, M., Pelegrin, A.J., Szareski, V.J., Meira, D., Meier, C. (2016). Genetic gain for cold tolerance among and within populations of palisade gras. Australian Journal of Basic and Applied Sciences, 10: 217-224.
Gehling, V.M., Pedó, T., Martinazzo, E.G., Aumonde, T.Z., Villela, F.A. (2018). Effect of soil water logging stress on the physiological performance of seeds and on the productivity of rye plants. Agrociência, 19: 41-47.
Gehlling, V.M., Segalin, S.R., Troyjack, C., Pimentel, J.R., Carvalho, I.R., Szareski, V.J., Dubal, I., Lautenchleger, F., Souza, V.Q., Schuch, L.O.B., Pedo, T., Villela, F.A., Aumonde, T.Z. (2015). Temporal Waterlogging and Physiological Performance of Wheat (Triticumaestivum L.) Seeds. Journal of Agricultural Science, 10: 363.
Koch, F., Monteiro, M.A., Pimentel, J.R., Troyjack, C., Dubal, I., Borges, E., Szareski, V.J., Demari, G., Jaques, L.A., Carvalho, I.R., Villela, F.A., Pedó, T., Aumonde, T.Z. (2018). Ecology of Upland Rice Plants and Seeds Subjected to Growth Regulator. American Journal of Plant Sciences, 60: 73-79.
Kumutha, D., Sairam, R.K., Ezhilmathi, K., Chinnusamy, V., Meena, R.C. (2008). Effect of waterlogging on carbohydrate metabolism in pigeon pea (Cajanus cajan L.): Upregulation of sucrose synthase and alcohol dehydrogenase. Plant Science, v. 175, p. 706-716.
Lopes, N.F., Lima, M.G.S. (2015). Fisiologia da produção. Viçosa. Ed. UFV, p.492.
Meira, D., Carvalho, I.R., Nardino, M., Follmann, D.N., Pelegrin, A.J., Szareski, V.J.,Ferrari, M., Olivoto, T., Meier, C., Souza, V.Q. (2016). Pathanalysis and dissimilarity in soybean with indeterminate habit. International Journal of Current Research, 8: 39568-39573.
Monteiro, M.A., Aisenberg, G.R., Koch, F., Salau, G.M., Behenck, J.P.O., Demari, G., Szareski, V.J., Carvalho, I.R., Nardino, M., Schuch, L.O.B, Pedó, T., Villela, F.A., Aumonde, T.Z. (2016). Yield and vigor of corn seeds under the influence of flooding periods. African Journal of Agricultural Research, 11: 3240-3245.
Nakagawa J (1999) Stability tests based on seedling performance.In: Kryzanowski FC, Vieira RD, França Neto JB. (Ed.). Vigor de sementes: conceitos e testes. Londrina: Abrates, 2: 1-24.
Nardino, M., Carvalho, I.R., Follmann, D.N., Pelegrin, A.J., Ferrari, M., Olivoto T., Szareski, V.J., Volpi, G.B., Souza, V.Q. (2016). Genetic breeding backround and sustainability in environments of production: A review. International Journal of Current Research, 8: 39629-39632.
Pedó, T., Koch, F., Delias, D.S., Aisenberg, G.R., Szareski, V.J., Carvalho, I.R., Nardino, M., Souz,a V.Q., Amarante, L., Villela, F.A., Aumonde, T.Z. (2016). Antioxidant enzyme activity and initial growth of wheat, rye and bean under soil flooding. International Journal of Current Research, 8: 36635-36642.
Perata, P., Armstrong, W. & Voesenek, L.A.C.J. (2011). Plants and flooding stress. New Phytologist. 190: 269-273.
Peske, S.T., Villela, F.A., Meneguello, G.E. (2012). Sementes: Fundamentos Científicos e Tecnológicos, 3 ed, 573p.
Rigo, G.A., Schuch, L.O.B., Barros, W.S., Vargas, R.L, Szareski, V.J., Carvalho, I.R., Pimentel, J.R., Troyjack, C., Jaques, L.B.A., Da,. Rosa, T.C., Souza, V.Q., Aumonde, T.Z., Pedó, T. (2018). Effects of Macronutrients in the Physiological Quality of Soybean Seeds. Journal of Agricultural Science. 10: 312-318.
Rosa, T.D., Pedó, T., Martinazzo, E.G., Gehling, V.M., Aisenberg, G.R., Aumonde, T.Z., Villela, F.A. (2015). Soil waterlogging: effect on the initial growth of oat (Avena sativa L.). Scientia Agraria Paranaensis, 14(2): 127-131.
Sachs, M., Vartapetian, B. (2007). Plant anaerobic stress I. Metabolic adaptation to oxygen deficiency. Plant Stress, 1(2): 123135.
Santos, E..L, Cattelan, A.J., Prete, C.E.C., Neumaier, N., Oliveira, M.C.N., Faria, J.R.B., Carvalho, J.F.C., Nepomuceno, A.L. (2012) Water stress affecting nodulation, oil, protein and grain yield of soybean cultivars. Global Science and Technology, 5(2): 109-120.
Shingaki-Wells, R.N., Huang, S., Taylor, N.L., Carroll, A.J., Zhou, W., Millar, A.H. (2011). Differential molecular responses of rice and wheat coleoptiles to anoxia reveal novel metabolic adaptations in amino acid metabolism for tissue tolerance. Plant Physiology, 156: 1706-1724.
Soares, V.N., Szareski, V.J., Demari, G.H., Martinazzo, E.G., Villela, F.A., Pedó, T., Carvalho, I.R., Nardino, M., Souza, V.Q., Aumonde, T.Z. (2016). Stress due to soil flooding and differential plysiological responses to initial growht of wheat plants. Australian Journal of Basic and Applied Sciences, 10: 260-266.
Szareski, V.J., Carvalho, I.R., Nardino, M., Pelegrin, A.J., Ferrari, M., Gaviraghi, R., Demari, G.H., Follmann, D.N., Warths, C.A., Souza, V.Q. (2016). Competition of soybean genotypes cultivated in lowlands of Rio Grande do Sul, Brazil. International Journal of Current Research, 8: 39714-39718.
Szareski, V.J., Carvalho, I.R., Kehl, K., Levien, A.M., Nardino, M., Dellagostin, S.M., Demari, G.H., Lautenchleger, F., Villela, F.A, Pedo, T., Souza, V.Q., Aumonde, T.Z. (2018). Evaluation of the adaptability and stability of wheat genotypes using a phenotypic index of seed vigor. Pesquisa Agropecuária Brasileira, 53: 727-735.
Troyjack, C., Pimentel, J.R., Dubal, I., Escalera, R.A.V., Jaques, L.A., Koch, F., Monteiro, M.A., Demari, G.H., Szareski, V.J., Carvalho, I.R., Schuch, L.O.B., Aumonde, T.Z., Pedó, T. (2018). Nitrogen fertilization on maize sowing: plant growth and seed vigor. American Journal of Plant Sciences, 2018: 83-97.
Vieira, Junior, P.A., Dourado, Neto. D. (2008). Milho. In: Castro PRC, Kluge RA, Sestari I. (Eds.) Manual de fisiologia vegetal: fisiologia de cultivos. 1ª ed. Piracicaba: Ceres: 130-156
Wang, X., Liu, T., Li, C., Chen, H. (2012). Effects of soil flooding on photosynthesis and growth of Zea mays L. seedlings under different light intensities. African Journal of Biotechnology, 11: 7676-7685.
Yemm, E.M., Cocking, E.C. (1955). Estimation of amino acids by ninhidrin. Analyst, 80: 209-213.
Zabalza, A., Dongen, J.T.V., Froehlich, A, Oliver, S.N., Faix, B., Gupta, K.J., Schmälzlin, E., Igal, M., Orcaray, L., Royuela, M., Geigenberger, P. (2008). Regulation of respiration and fermentation to control the plant internal oxygen concentration. Plant Physiology, Rockville, 149(2): 1087-1098.
Zanatta, E., Szareski, V.J., Carvalho, I.R., Pimentel, J.R., Troyjack, C., Dellagostin, S.M., Demari, G., Lautenchleger, F., Souza, V.Q., MartinazzoEG,Villela FA, Aumonde TZ (2018). Pre-harvest Desiccation: Productivity and Physical and Physiological Inferences on Soybean Seeds During Storage. Journal of Agricultural Science, 10: 354.
Ziegler, P. (1995). Carbohydrate degradation during germination. In: Kigel, J.;Galili, G. (Ed.). Seed development and germination. New York: Marcel Dekker: 447-474.
Zimmer, G., Koch, F., Carvalho, I.R., Szareski, V.J., Demari, G.H., Nardino, M., Follmann, D,N., Souza, V.Q., Aumonde, T.Z., Pedó, T. (2016). Seed quality and initial performance os seedlings of soybean produced off-season in Rio Grande do Sul, Brasil. International Journal of Current Research, 8: 40325-403.
Author notes
angel.celente10@gmail.com

