

Asociaciones entre caracteres vinculados al crecimiento inicial en poblaciones espontáneas de Stapfochloa berroi de la Pampa Deprimida (Buenos Aires, Argentina)
Revista de la Facultad de Agronomía
Universidad Nacional de La Plata, Argentina
ISSN: 1669-9513
Periodicidad: Frecuencia continua
vol. 122, 125, 2023
Recepción: 29 Agosto 2022
Aprobación: 27 Septiembre 2022
Resumen: Stapfochloa berroi (Arechav.) P.M. Peterson es una gramínea perenne de crecimiento primavero-estival, nativa de Sudamérica, que en la Pampa Deprimida (Buenos Aires, Argentina) es clave debido a su valor forrajero y adaptación a ambientes que son restrictivos para otras especies forrajeras exóticas. Reincorporar germoplasma nativo seleccionado sería importante tanto para aumentar la productividad ganadera como para mantener y/o recuperar la biodiversidad en estos agroecosistemas. El objetivo fue estudiar asociaciones y relaciones funcionales entre algunos de los caracteres que podrían explicar el éxito de implantación en dos poblaciones espontáneas de S. berroi provenientes de estepas de halófitas de la Pampa Deprimida, evaluadas en dos sustratos. Se registró el peso individual de semillas para ambas poblaciones. Posteriormente, las semillas con su peso individual identificado se sembraron en bandejas plantineras con sustrato limitante (suelo de bajo alcalino) y sin limitantes (suelo de horizonte A de un argiudol típico) en invernáculo. A los 54 días, para cada individuo, se determinó: altura, longitud de la raíz adventicia más larga y total, número de raíces adventicias, de hojas verdes desplegadas y de macollas. También se determinó peso seco aéreo, radical y total. Se analizaron correlaciones (coeficiente de Pearson) y regresiones múltiples entre los caracteres. Para ambas poblaciones, en cada uno de los sustratos, se encontraron patrones de correlaciones similares y significativos (p≤0,05). Algunos caracteres de fácil medición (e.g. altura de planta, número de hojas) resultaron determinantes del peso seco total. Esto permitirá diseñar estrategias de selección indirecta independientemente del sustrato utilizado.
Palabras clave: mejoramiento genético, forrajeras nativas, regresión, correlación, restauración, pastizales naturales.
Abstract: Stapfochloa berroi (Arechav.) P.M. Peterson is a spring-summer growing perennial grass, native to South America, which in the Pampa Deprimida (Buenos Aires, Argentina) is key due to its forage value and adaptation to environments that are restrictive for other exotic forage species. Reincorporating selected native germplasm would be important both to increase livestock productivity and to maintain and/or recover biodiversity in these agroecosystems. The objective was to study associations and functional relationships among some of the characters that could explain the success of implantation in two spontaneous populations of S. berroi from halophyte steppes of the Pampa Deprimida, evaluated in two substrates. Individual seed weight was recorded for both populations. Subsequently, seeds with their individual weights identified were sown in seedling trays with limiting substrate (low alkaline soil) and with not-limiting substrate (horizon A from a typical argiudol soil) in a greenhouse. After 54 days, for each individual, the following were determined: height, total plant length and length of the longest adventitious root, number of adventitious roots, number of green leaves unfolded and number of tillers. Aerial, root and total dry weight were also determined. Correlations (Pearson's coefficient) and multiple regressions between characters were analyzed. For both populations, in each of the substrates, similar and significant (p≤0.05) correlation patterns were found. Some easily measured traits (e.g. plant height, number of leaves) were found to be determinants of total dry weight. This will allow to design strategies of indirect selection independently of the substrate used.
Keywords: plant breeding, forage native species, regression, correlation, restoration, natural grasslands.
INTRODUCCIÓN
La Pampa Deprimida ocupa aproximadamente 9 millones de hectáreas en la provincia de Buenos Aires (Argentina) al este del sistema de sierras de Tandilia e incluye a las cuencas de Laprida y del Río Salado, que representan el 25 y el 75% de la superficie de la región respectivamente (Cid et al., 2011). La actividad productiva principal es la cría de ganado bovino (Soriano et al., 1991), siendo el pastizal natural la forma de vegetación predominante de la región (Batista et al., 2005) y el recurso forrajero que sostiene la actividad ganadera (Faverin & Machado, 2019).
La vegetación de una región está constituida por el mosaico de comunidades vegetales que pueblan los distintos paisajes que la definen. Si bien, actualmente, en la región Pampeana los agroecosistemas han reemplazado al pastizal natural y sólo en áreas con impedimentos serios para la agricultura se conservan pastizales seminaturales extensos (León & Burkart, 1998), Burkart et al. (2005) describen cuatro comunidades de pastizales que se diferencian claramente entre ellas y están adaptadas a diferentes condiciones edafoclimáticas: i) la pradera de mesófitas, en suelos Argiudoles o Hapludoles que ocupan las posiciones más altas del relieve; ii) la pradera húmeda de mesófitas, que ocupa áreas planas y depresiones ligeras, con algunas limitaciones de drenaje y rasgos de sodicidad subsuperficial; iii) la pradera de hidrófitas, en suelos Argiacuoles, Argialboles, Cromuderts acuenticos, Calciacuoles y Udifluventes, con severos problemas de drenaje pero sin problemas de sodicidad; y iv) la estepa de halófitas, en suelos Alfisoles (Natracualfes, Natrudalfes, Fragiacualfes, Natrustalfes, entre otros), con deficiencias en el drenaje y altas concentraciones de sales y sodio desde superficie, ambientes conocidos como “bajos alcalinos”. El 24% de la superficie de la provincia de Buenos Aires (7,08 millones de hectáreas) está ocupada por suelos hidrohalomórficos (Natracuoles, Natracualfes, etc.) donde al menos el 68% de los mismos (4,75 millones de hectáreas) se hallan en la Pampa Deprimida (Miaczynski, 1995), dando una idea de cuán relevantes son estos ambientes para la producción ganadera.
La estepa de halófitas se caracteriza por la poca cobertura vegetal y por estar compuesta por especies de limitado valor forrajero que requieren un manejo cuidadoso del pastoreo para evitar su disminución y eventual desaparición de la comunidad (Burkart et al., 2005). Desde el punto de vista forrajero muestra un crecimiento del pastizal marcadamente primavero-estival, con un promedio de productividad primaria neta aérea que ronda los 2500 kg de materia seca por ha/año. Por las condiciones fisicoquímicas de sus suelos, las implantaciones de pasturas con mezclas de especies forrajeras templadas no resultan siempre satisfactorias en términos productivos y económicos (Rossi, 2008). Estos agroecosistemas, ya de por sí frágiles como consecuencia de las severas limitaciones edafoclimáticas imperantes, se vuelven más vulnerables por efecto del pastoreo de grandes herbívoros, que afectan principalmente la composición de especies al provocar daños selectivos en plantas individuales por herbivoría y pisoteo (Vecchio, 2014). Bajo pastoreo continuo la comunidad está dominada por Distichlis scoparia (Nees ex Kunth) Arechav. y Distichis spicata (L) Green (León et al., 1979), especies forrajeras fibrosas, con baja digestibilidad (Escobar Hernández et al., 2005).
Una estrategia válida para mejorar estos ambientes es el mejoramiento genético de especies forrajeras nativas de esta región para su reincorporación a los pastizales. Según Pistorale et al. (2008) las especies forrajeras nativas y/o naturalizadas se encuentran adaptadas a las condiciones específicas del ambiente; así, según Mujica (2010a; b), la mejora genética debería orientarse a lograr la combinación de los efectos de la selección natural en el ambiente local y los de la selección artificial para caracteres de interés agronómico. Existen diversos antecedentes en la identificación de especies vegetales nativas o naturalizadas y adaptadas a condiciones edafoclimáticas limitantes que han sido incluidas y domesticadas en programas de mejora genética que tuvieron el objetivo de seleccionar germoplasma para reincorporar a estos ambientes degradados, tanto para aumentar la biodiversidad como para mejorar la productividad primaria y secundaria, la cobertura del suelo y favorecer los procesos ecosistémicos. Esta estrategia de mejora genética se ha aplicado, por ejemplo, a Elymus scabrifolius (Döll) J. H. Hunz. (agropiro criollo) (UNL Noticias, 2004), Trichloris crinita (Lag.) Parodi (plumerillo) (INTA informa, 2013), y Lotus tenuis Mill (Mujica, 2014).
Stapfochloa berroi (Arechav.) P.M. Peterson (S. berroi, syn. Chloris berroi) es una gramínea perenne de crecimiento primavero-estival y buen valor forrajero, nativa de Sudamérica -sur de Brasil, Uruguay y centro y norte de Argentina (Anderson, 1974; Royal Botanic Gardens) que se encuentra en pastizales importantes para la producción animal. Específicamente, en Argentina, se halla en los pastizales del norte de la Pampa Deprimida y tiene la particularidad de encontrarse casi exclusivamente en los ambientes halomórficos (León et al., 1979). Bajo pastoreo rotativo representa un alto porcentaje de la cobertura total de la comunidad de las estepas de halófitas a fines de verano y principios de otoño; por el contrario, bajo pastoreo continuo la densidad y la cobertura de la especie disminuyen significativamente (Ansin et al., 2005; Vecchio et al., 2006; 2019). Debido a su buen valor forrajero y a su adaptación a estos ambientes restrictivos (suelos sódicos, bajo contenido de materia orgánica, anegamiento, sequía) para otras especies forrajeras exóticas, S. berroi resulta un recurso clave para la región.
Desde el punto de vista del mejoramiento genético, además de realizar estudios de variabilidad de los caracteres de interés, es importante poder estudiar sus posibles asociaciones y/o relaciones funcionales. Éstas son relevantes ya que posibilitarán conocer si la selección por un carácter afectará a otros caracteres correlacionados. Según Falconer & Mackay (1996) esto se denomina respuesta correlacionada y es útil para seleccionar un carácter primario de difícil determinación a través de uno secundario de medición más sencilla. Esta respuesta correlacionada a la selección puede ser deseable cuando dos caracteres están asociados de manera favorable, así la selección por un primer carácter lograría mejorar a un segundo carácter. Contrariamente, puede ocurrir que cuando se selecciona sobre las poblaciones para un determinado carácter se arrastre involuntariamente a otro, normalmente no deseado (Cubero, 2003). En cuanto a las relaciones funcionales entre caracteres, su utilidad radica en permitir establecer relaciones causa-efecto entre caracteres complejos (e.g. crecimiento o vigor inicial de planta) y caracteres más sencillos (e.g. altura de planta, número de hojas). El vigor de planta o plántula (según el estadio en que se la estudie), o crecimiento inicial, se define generalmente por el peso de la misma luego de un periodo de crecimiento (Smart et al., 2003) en un ambiente determinado (Qualls et al., 1968). Existen numerosos antecedentes en el estudio de las asociaciones entre caracteres en especies vegetales de uso forrajero, como, por ejemplo, Panicum maximum Jacq. (Seguí et al., 1989), Paspalum dilatatum Poir (pasto miel) (García et al, 2000), Pappophorum vaginatum Buckley (Entio, 2014), Phalaris aquatica L. (falaris bulbosa) (Spara et al., 2014), y Schedonorus phoenix (Scop.) Holub (festuca alta) (Buratovich, 2015).
En el caso de S. berroi, dada la escasez de información disponible sobre sus características y su importancia como componente de las estepas de halófitas de los bajos alcalinos de la Pampa Deprimida, resulta importante generar conocimiento sobre posibles asociaciones y relaciones funcionales entre caracteres vinculados al control de la implantación, como el crecimiento o vigor inicial de planta, y sus posibles determinantes y/o componentes (e.g. altura de planta, número de hojas, número de macollas).
La hipótesis de este estudio es que existen correlaciones y relaciones funcionales entre ciertos caracteres simples (e.g. número de hojas, macollas y raíces adventicias, altura de planta, etc.) y el carácter complejo crecimiento o vigor inicial de las plantas (i.e. acumulación de peso seco) en poblaciones espontáneas de S. berroi. El objetivo fue estudiar posibles asociaciones y relaciones funcionales de caracteres vinculados al crecimiento o vigor inicial de planta en dos poblaciones espontáneas de S. berroi provenientes de la Pampa Deprimida (Buenos Aires, Argentina) cultivadas en dos sustratos de características físicoquímicas contrastantes, uno extraído del ambiente de origen de la especie (natracualf) y el otro obtenido de un ambiente de mejor calidad edáfica (argiudol típico).
MATERIALES Y MÉTODOS
RECOLECCIÓN DE LAS SEMILLAS
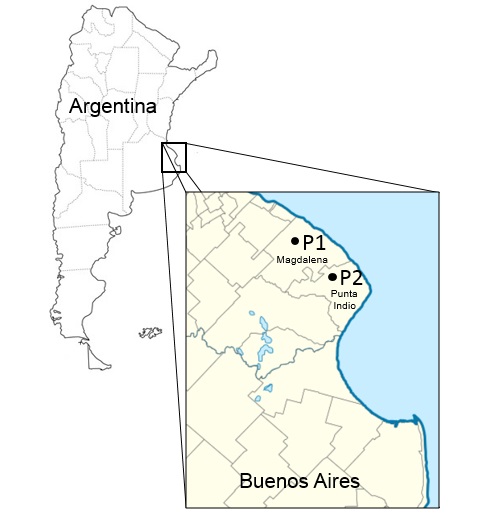
Se recolectaron semillas de dos poblaciones espontáneas (P) de S. berroi en estepas de halófitas de la Pampa Deprimida, provincia de Buenos Aires, en los partidos de Magdalena (P1: 35° 16´ 24,67"S; 57° 38´ 5,14"O) y Punta Indio (P2: 35º 27´S; 57º 22´O) en enero de 2016 (Figura 1). Las recolecciones fueron en áreas al azar de 100 m., separadas por un mínimo de 10 km (ENSCONET, 2009). Cada área se consideró como una población diferente. Se tomaron en una misma bolsa un mínimo de 50 inflorescencias de 50 individuos diferentes por área (Marshall & Brown, 1975). Las inflorescencias se cosecharon en el estadio fenológico de madurez/desprendimiento de semillas (color amarillo pajizo). Luego de la recolección y hasta el momento del ensayo estas semillas estuvieron almacenadas en bolsas de papel madera rotuladas y en heladera a 4 ºC.
En Magdalena la precipitación media anual histórica es de 1018 mm (1980 a 2016). La temperatura generalmente varía de 6 °C a 27 °C a lo largo del año, siendo las máximas y mínimas medias de 27 °C (enero) y 6 °C (junio y julio), respectivamente (Weatherspark, 2016a). En Verónica, ciudad cabecera del partido de Punta Indio, la precipitación media anual histórica es de 958 mm (1980 a 2016). La temperatura generalmente varía de 6 °C a 26 °C a lo largo del año, siendo las máximas y mínimas medias de 26 °C (enero) y 6 °C (julio), respectivamente (Weatherspark, 2016b).
PESO DE SEMILLAS, CULTIVO Y PROCESAMIENTO DE PLANTAS
Previo al ensayo se registró el peso individual de 400 cariópsides por población (PC) con sus coberturas (lemma y pálea) en balanza analítica (Mettler H54AR). Cada cariópside pesada se guardó envuelta individualmente en un sobre de papel absorbente rotulado. El 8 de octubre de 2019 se sembraron las semillas (cariópsides con sus envolturas) en bandejas plásticas de 30 celdas (15,5 cm altura, 27 cm ancho, 32 cm de largo y volumen de 160 cm. por celda), algunas de las cuales contenían sustrato extraído del horizonte A de un suelo argiudol típico (S1, sin limitantes), mientras que las restantes bandejas contenían sustrato extraído del horizonte superficial de un suelo natracualf de un bajo alcalino (S2, con limitantes) (Tabla 1). Ambos sustratos fueron tamizados previo al llenado las bandejas.
La siembra se realizó manualmente a una profundidad de 5 mm a razón de 2 semillas por celda, formando una recta horizontal en el centro de la misma, en la que cada extremo correspondió a una semilla numerada determinada.
Luego de la siembra las bandejas plantineras se llevaron a invernáculo. La temperatura (°C) y la humedad relativa (%) se registraron diariamente con un termohigrómetro digital modelo ThermoMeter TA318. El rango de temperaturas en el invernáculo durante el desarrollo del estudio fluctuó entre 41,5 °C y 11 °C y las temperaturas medias máximas y mínimas ± error estándar fueron 33,1 °C ± 0,74 y 17,1 °C ± 0,48, respectivamente. La humedad relativa fluctuó entre 100% y 22% y la humedad relativa media máxima y mínima ± error estándar fueron 84,2% ± 1,12 y 43,5% ± 2,36, respectivamente. El riego de las bandejas también se realizó diariamente hasta que comenzó a salir agua gravitacional por el fondo de la celda. Durante el cultivo se fueron eliminando plántulas de especies espontáneas pertenecientes al banco de propágulos de los sustratos. Además, alcanzado el estadio fenológico de segunda hoja expandida, se realizó un raleo al azar de las plantas para evitar la interferencia por competencia, procurando dejar una sola por celda.
A los 54 días desde la siembra las plantas se extrajeron de las celdas con su bloque de sustrato y se lavaron con agua corriente en bateas plásticas para eliminar el sustrato adherido. Las plantas lavadas se secaron con papel absorbente, se guardaron individualmente en bolsitas plásticas rotuladas y se conservaron en heladera hasta el momento de la evaluación de los caracteres de planta.
Para cada planta se determinó: altura (A), medida desde el cuello de la raíz hasta el extremo de la hoja más larga; longitud de la raíz seminal más larga (LR), medida desde el cuello de la raíz hasta el extremo de dicha raíz seminal; longitud total (LT= A+LR); longitud de la raíz adventicia más larga (Ladv); número de raíces adventicias (n°adv); número de hojas verdes desplegadas (H) y número de macollas (n°mac). Luego se disectó cada planta a la altura del cuello de la raíz, se llevaron a estufa a 60 °C durante 72 h, se determinó peso seco aéreo (PSA) y radical (PSR), y se calculó el peso seco total (PST= PSA+PSR). También se calcularon las relaciones PSA/PSR y A/LR.
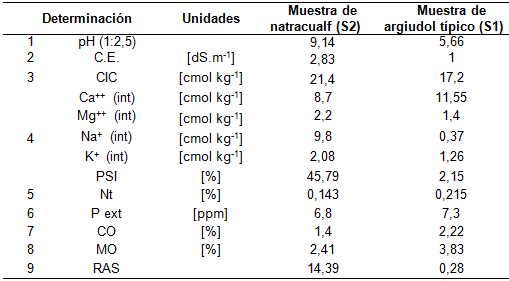
Características físico-químicas de los sustratos utilizados en el ensayo. Referencias: 1. pH, relación suelo / agua, 1:2,5, determinación potenciométrica; 2. Conductividad eléctrica, medida en el extracto de saturación, determinación conductimétrico; 3. Capacidad de intercambio catiónico, determinada por el método acetato de amonio 1N pH 7; 4. Cationes intercambiables, determinación de Ca++ y Mg++ por quelatometría con EDTA; Na+ y K+ por fotometría de llama; 5. Nitrógeno total, digestión húmeda, evaluación por método Microkjeldahl; 6. Fósforo extractable, método Bray Kurtz N°1; 7. Carbono fácilmente oxidable, método de Walkley – Black; 8. Materia orgánica. MO (%) = CO x 1,724; 9. RAS: Relación de adsorción de sodio.
ANÁLISIS ESTADÍSTICO
Se analizaron correlaciones entre caracteres (coeficiente de Pearson) y las relaciones funcionales de los caracteres sobre el PST mediante regresión múltiple (proceso de selección paso a paso hacia adelante). Todos los análisis estadísticos se realizaron con el software Infostat versión 2012 (Di Rienzo et al., 2012).
RESULTADOS
Los resultados del análisis de correlación de Pearson entre los caracteres estudiados mostraron que: I) en S1 ambas poblaciones presentaron una correlación significativa y positiva entre el PST y A, LT, A/LR, n°adv, H, n°mac y PC. Además, P1S1 mostró una correlación significativa aunque negativa entre PST y LR; II) en S2 el patrón de correlaciones fue similar a S1, las diferencias fueron que PST tuvo una correlación positiva y significativa con Ladv, aunque no presentó correlación con A/LR. Además, P1 tuvo una correlación significativa y positiva con LR (Tabla 2).
El análisis de regresión múltiple (proceso de selección paso a paso hacia adelante) de los caracteres estudiados sobre el PST fue significativo (p≤0,05) en ambas poblaciones. Se encontró que, para la combinación P1S1, con la inclusión de la altura de planta, el número de hojas y el número de raíces adventicias, se explicó un 72,31% de la variación del PST (p≤0,05); para la combinación P1S2, los caracteres número de hojas, número de raíces adventicias y longitud total de planta explicaron el 69,13% de la variación del PST (p≤0,05); para la combinación P2S1, los caracteres altura de planta, número de raíces adventicias y número de macollas explicaron el 75,01% de la variación del PST (p≤0,05); y, para la combinación P2S2, los caracteres altura de planta, número de hojas, número de raíces adventicias y longitud total explicaron el 75% de la variación del PST (p≤0,05) (Tabla 3).

Correlación de caracteres vinculados al crecimiento o vigor inicial de planta (PST) en dos poblaciones (P1 y P2) de S. berroi para dos sustratos (S1 y S2) (n=80). Coeficiente de correlación de Pearson. ns: no significativo; *: p≤0,05; **: p≤0,01. Referencias: PST: peso seco total; A: altura de planta; LR: longitud radical; LT: longitud total; Ladv: longitud de la raíz adventicia más larga; nºadv: número de raíces adventicias; nºmac: número de macollas; H: número de hojas; PC: peso de cariópside.
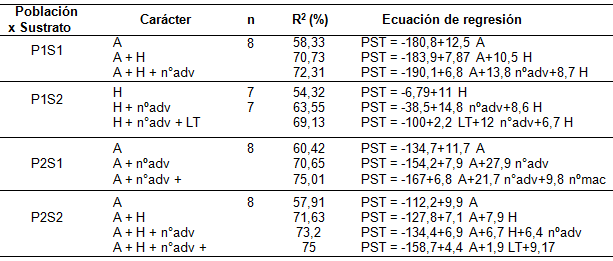
Análisis de la regresión múltiple (proceso de selección paso a paso hacia adelante) de los caracteres evaluados sobre el peso seco total por planta (PST) en dos poblaciones de S. berroi (P1 y P2) en dos sustratos (S1 y S2). Referencias: PST: peso seco total; A: altura de planta; LT: longitud total; nºadv: número de raíces adventicias; nºmac: número de macollas; H: número de hojas.
DISCUSIÓN Y CONCLUSIONES
Este estudio tuvo como objetivo determinar posibles asociaciones y relaciones funcionales entre algunos caracteres clave para el crecimiento o vigor inicial de planta, que podrían contribuir al éxito de la implantación, en dos poblaciones de S. berroi recolectadas en estepas de halófitas de la Pampa Deprimida (provincia de Buenos Aires, Argentina). Ambas poblaciones se evaluaron en un sustrato similar al de origen (S2: suelo del horizonte superficial de un bajo alcalino de la Pampa Deprimida) y en un sustrato sin limitantes físicoquímicas (S1: suelo del horizonte A de un argiudol típico). De esta manera se buscó estudiar los patrones de correlaciones y relaciones funcionales para ambas poblaciones y evaluar si resultaban similares en ambos sustratos, tanto para conocer la posible utilización indistinta de uno u otro sustrato en futuros procesos de selección como para identificar caracteres asociados y/o determinantes del crecimiento o vigor inicial de las plantas y orientar estrategias de selección indirecta en posibles planes de mejora de la implantación de la especie.
Se observó que el patrón de correlaciones fue similar para ambas poblaciones y sustratos. El estudio de las respuestas correlacionadas sugiere que se podría obtener un progreso genético más rápido seleccionando un carácter correlacionado (o secundario) en vez del carácter de interés (o primario), especialmente cuando el carácter primario es difícil o costoso de medir (Falconer & Mackay, 1996). Así, de modo general, se observó que la altura de planta, la longitud total, el número de raíces adventicias, el número de hojas, el número de macollas y el peso de la cariópside se correlacionaron de manera positiva y significativa con el peso seco total de las plantas. El peso de la cariópside se correlacionó significativamente (p≤0,05) con el peso seco total de las plantas, aunque con bajos valores de coeficiente de correlación. En el caso de P2S1, esta correlación, además de ser significativa (p≤0,05), resultó negativa. Esto indicaría que, así como lo observado en otras especies, el peso de la cariópside podría utilizarse como predictor del vigor de plántula. De esta forma, lo hallado en S. berroi coincide con lo obtenido para leguminosas forrajeras como alfalfa (Beveridge & Wilsie, 1959) y Lotus L. (Collado & Mujica, 2002) y para gramíneas como Lolium perenne L. (Thomas, 1966), Andropogon hallii Hack (Glewen & Vogel, 1984), Panicum virgatum L. (Smart & Moser, 1999); aunque difiere de lo obtenido para Pappophorum vaginatum Buckley (Entio et al., 2014). La similitud en los patrones de correlaciones hallados para ambas poblaciones entre el vigor de planta (peso seco total) y otros caracteres vinculados al mismo de fácil medición (e.g. altura de planta, número de macollas, número de hojas) en los dos sustratos para las condiciones ensayadas, permitirían realizar un procedimiento de selección indirecta independientemente de si se recolecta sustrato in-situ de los bajos alcalinos o si se utiliza sustrato extraído del horizonte A de un argiudol típico. Sin embargo, en futuros trabajos deberían estudiarse las correlaciones genéticas entre los caracteres abordados.
Mientras que en el análisis de correlación se mide el nivel de asociación entre variables sin unidades físicas y sin especificar cuál es la causa y cuál la consecuencia, el análisis de regresión tiene unidades físicas e implica relación causa-efecto (Gomez & Gomez, 1984). Del análisis de regresión múltiple resultó que el peso seco total (PST) estuvo explicado en mayor o menor medida por diferentes caracteres dependiendo del tratamiento. Los caracteres aéreos fueron importantes en la determinación del PST, dado que definen su influencia sobre la capacidad de producción de fotoasimilados de la planta. La altura de planta es un carácter fácilmente medible a campo y explicó el 58,33%, 60,42% y 57,91% del PST en las combinaciones P1S1, P2S1 y P2S2, respectivamente; por lo que se constituye como un importante carácter de selección indirecta del peso seco total inicial de las plantas. La relación funcional encontrada entre la altura de planta y el peso seco total fue también observada por Entio et al. (2014) para poblaciones de P. vaginatum y por Ré et al. (2006) para poblaciones de Bromus auleticus Trin ex Nees (cebadilla chaqueña). El número de raíces adventicias fue un carácter relevante para el peso seco total en los cuatro tratamientos, pudiéndose asociar a que el mismo se corresponde con una mayor captación de agua y nutrientes del suelo (Wilson et al., 1976); sin embargo, su observación no puede realizase a simple vista y requiere de la extracción de la planta del suelo, por lo que sería poco práctica su utilización en una selección indirecta. También se observó que el número de hojas explicó parte de la variación del peso seco total en las combinaciones P1S1, P1S2 y P2S2. Para P2, en el sustrato sin limitantes (S1), el número de macollas representó un 5% de la variación del peso seco total. Tanto el número de hojas como el número de macollas son fácilmente observables y medibles a campo, aunque su menor incidencia en la determinación del peso seco total en comparación a la altura de planta hace que sea menos conveniente su utilización como estrategia de selección indirecta. Esto difiere de lo observado por Entio et al. (2014) para dos poblaciones de P. vaginatum, en las que la altura de planta y el número de macollas explicaron hasta el 78% del peso seco total. Del análisis de regresión se observa que, de acuerdo con la población y sustrato, la altura de planta o el número de hojas explican más del 54% de la variabilidad para el crecimiento o vigor inicial de planta, pudiendo ser considerados como determinantes de dicho carácter y utilizados como predictores del mismo independientemente del sustrato utilizado.
BIBLIOGRAFÍA
Anderson, D.E. (1974). Taxonomy of the genus Chloris (Gramineae). Brigham Young University Science Bulletin. Biological Series, 19(2),1.
Ansin, O.E., Oyhamburu, E.M., Vecchio, M.C., Cordero, M.I. y Heguy, B. (2005). Estructura y condición forrajera de un pastizal alcalino ante diferentes manejos de pastoreo. III Congreso Nacional de manejo de pastizales. Rosario, Argentina.
Batista, W.B., Taboada, M.A., Lavado, R.S., Perelman, S.B. y León, R.J. (2005). Asociación entre comunidades vegetales y suelos en el pastizal de la Pampa Deprimida. En M. Oesterheld, M.R. Aguiar, C.M. Ghersa y J.M. Paruelo (coords.), La heterogeneidad de la vegetación de los agroecosistemas, Un homenaje a Rolando León. (pp.113-129). Editorial Facultad de Agronomía.
Beveridge, J.L. y Wilsie, C.P. (1959). Influence of depth of planting, seed size, and variety on emergence and seeding vigor in Alfalfa. Agronomy Journal, 51(12), 731-734.
Buratovich, M.V. (2015). Correlaciones genotípicas de variables morfogenéticas y estructurales con el rendimiento de forraje en festuca alta Schedonorus phoenix (Scop.) Holub. [Tesis de grado]. Escuela de Ciencias Agrarias, Naturales y Ambientales, Universidad Nacional del Noroeste de la Provincia de Buenos Aires.
Burkart, S.E., Garbulsky, M.F., Ghersa, C.M., Guerschman, J.P., León, R.J.C., Oesterheld, M., Paruelo, J.M. y Perelman, S.B. (2005). Las comunidades potenciales del pastizal pampeano bonaerense. En M. Oesterheld, M.R. Aguiar, C.M. Ghersa y J.M. Paruelo (coords.), La heterogeneidad de la vegetación de los agroecosistemas, Un homenaje a Rolando León. (pp 379-395). Editorial Facultad de Agronomía.
Cid, M.S., Grecco, R.F., Oesterheld, M., Paruelo, J.M., Cibils, A.F. y Brizuela, M.A. (2011). Grass-fed beef production systems of Argentina's flooding pampas. Understanding ecosystem heterogeneity to improve livestock production. Outlook on Agriculture, 40(2),181-189. https://doi.org/10.5367/oa.2011.0040
Collado, M.B. y Mujica, M.M. (2002). Aprovechamiento de la heterosis y rol de dos caracteres determinantes del vigor de las plántulas de Lotus glaber. XXI Congreso Argentino de Genética, Corrientes, Argentina.
Cubero, J.I. (2003). Introducción a la Mejora Genética Vegetal. (2ª ed.). Editorial Mundi Prensa.
Di Rienzo, J.A., Casanoves, F., Balzarini, M.G., Gonzalez, L., Tablada, M. y Robledo, C.W. (2012). InfoStat versión 2012. Grupo InfoStat, Facultad de Ciencias Agrarias, Universidad Nacional de Córdoba, Argentina.
Entio, L.J., Mujica, M.M., Busso, C., Torres, Y., Montenegro, O., Ithurrart, L., Giorgetti, H., Rodríguez, G., Bentivegna, D., Brevedan, R., Fernández, O., Baioni, S., Fioretti, M. y Tucat, G. (2014). Variabilidad y correlaciones de caracteres vinculados al vigor de plántula en dos poblaciones naturales de Pappophorum vaginatum Buckley. Revista de la Facultad de Ciencias Agrarias Universidad Nacional de Cuyo, 46(2), 223-230.
Escobar-Hernández, A., Troyo-Diéguez, E., García-Hernández, J.L., Hernández-Contreras, H., Murillo-Amador, B., & López-Aguilar, R. (2005). Potencial forrajero del pasto salado Distichlisspicata L. (Greene) en ecosistemas costeros de Baja California Sur, México por el método de “componentes principales”. Técnica pecuaria en México, 43(1), 13-25.
ENSCONET. (2009). Manual para la Recolección de Semillas de Especies Silvestres. Royal Botanic Gardens & Universidad Politécnica de Madrid.
Falconer, D.S. (1970). Introducción a la genética cuantitativa. Editorial CECSA.
Falconer, D.S. y Mackay, T.F.C. (1996). An introduction to quantitative genetics. (4ta ed.) Longman, Scientific & Technical.
Faverin, C. y Machado, C. (2019). Tipologías y caracterización de sistemas de cría bovina de la Pampa Deprimida. Chilean Journal of Agricultural & Animal Sciences, 35(1), 3-13.
García, M.V., Arturi, M.J. y Ansín, O.E. (2000). Potencial de mejora en caracteres cuantitativos de Paspalum dilatatum Poir. en poblaciones del noreste bonaerense. Revista de la Facultad de Agronomía, 104(2), 105-111.
Glewen, K.L. y Vogel, K.P. (1984). Partitioning the genetic variability for seedling growth in sand bluestem into its seed size and seedling vigor components. Crop Science, 24, 137-141.
Gomez, K. y Gomez, A. (1984). Statistical procedures for agricultural research with emphasis on rice. (2da ed.). John Willey and Sons.
INTA informa. (2013) Una forrajera promisoria para zonas marginales. Instituto Nacional de Tecnología Agropecuaria.
León, R.J.C. y Burkart, S. (1998). El pastizal de la Pampa Deprimida: Estados alternativos. Ecotrópico. Revista de la Sociedad Venezolana de Ecología, 11(2), 121-130.
León, R.J.C., Burkart, S. y Movia, C.P. (1979). Relevamiento fitosociológico del pastizal del Norte de la Depresión del Salado. Serie Fitogeográfica. INTA Buenos Aires.
Marshall, D.R. y Brown, A.H.D. (1975). Optimum sampling strategies in genetic conservation. En: Frankel, O.H. y Hawkes, J.G. (Eds.). Crop genetic resources for today and tomorrow. (pp. 53-80). Cambridge University Press.
Miaczynski, C. (1995). Los suelos hidromórficos e hidrohalomórficas de la provincia de Buenos Aires. Revista de la Facultad de Agronomía, 15, 23-36.
Mujica, M.M. (2010a). Estrategias de selección y rol de la mejora genética de especies nativas y naturalizadas para una ganadería pastoril sustentable. Jornadas de Mejoramiento Genético de Forrajeras, La Plata, Argentina.
Mujica, M.M. (2010b). Estrategias y resultados de una experiencia de investigación tecnológica y mejoramiento genético en Lotus tenuis. V Taller Interdisciplinario de Lotus: Aspectos genéticos, Moleculares y Ecofisiológicos de Lotus spp y sus simbiontes, INTECH, Chascomús, Argentina.
Mujica, M.M. (2014). 1ra Exposición de productos del Mejoramiento Genético Vegetal generados en la FCAyF-UNLP.
Pistorale, S.M., Abbott, L.A. y Andrés, A. (2008). Diversidad genética y heredabilidad en sentido amplio en agropiro alargado, Thinopyrum ponticum. Ciencia e Investigación Agraria, 35(3), 259-264.
Qualls, M. y Cooper, C.S. (1968). Germination, growth and respiration rates of birdsfoot trefoil at three temperatures during the early non-photosynthetic stage of development. Crop Science, 8, 758-760.
Ré, A.E., De Battista, J.P. y Costa, M. (2006). Variabilidad de caracteres asociados al vigor de plántula en cebadilla chaqueña. 2. Correlaciones genéticas, coheredabilidad y respuesta indirecta a la selección. Revista Argentina de Producción Animal, 26(1),121-262.
Rossi, C. (2008). Mejoramiento de la Condición Forrajera en Bajos Alcalino-Sódicos con Especies Subtropicales en la Cuenca del Salado. Sitio Argentino de Producción Animal.
Royal Botanic Gardens (s/f). Stapfochloa berroi. Royal Botanic Gardens, Plants of the World Online.
Seguí, E., Tomeu, A. y Machado, H. (1989). Asociaciones entre caracteres individuales y su importancia en el mejoramiento genético de la especie Panicum maximum Jacq. Pastos y Forrajes,12, 219-226.
Smart, A.J. y Moser, L.E. (1999). Switchgrass seedling development as affected by seed size. Agronomy Journal, 91, 335-338.
Smart, A.J., Moser, L.E. y Vogel, K.P. (2003). Establishment and Seedling Growth of Big Bluestem and Switchgrass Populations Divergently Selected for Seedling Tiller Number. Crop Science, 43, 1434-1440.
Soriano, A., León, R.J.C., Lavado, R.S., Deregibus, V.A, Cahuepé, M.A., Scaglia, O.A., Velázquez, C.A. y Lemcoff, J.H. (1991). Río de La Plata grassland. En Coupland, R.T. (Ed.), Ecosystem of the World- Natural grassland (pp. 367-407). Elsevier Science Publishers.
Spara, F., Abbott, L., Wolff, R. y Vernengo, E. (2014). Estimación de parámetros genéticos y su utilidad para la selección por producción de semilla en Phalaris aquatica L. Journal of Basic & Applied Genetics, 25, 31-40.
Thomas, R.L. (1966). The influence of seed weight on seedling vigour in Lolium perenne. Annals of Botany, 30, 111-121.
UNL Noticias. (2004). Científicos logran mejorar genéticamente una forrajera nativa. https://www.unl.edu.ar/
Vecchio, M.C. (2014). Modificaciones en la vegetación y el suelo inducidos por el manejo del pastoreo en la estepa de halófitas de la Pampa Deprimida. [Tesis Magister]. Facultad de Agronomía, Universidad de Buenos Aires.
Vecchio, M.C., Heguy, B., Oyhamburu, E.M., Cordero, M.I. y Ansin, O.E. (2006). Cómo influyen las distintas frecuencias de pastoreo a las comunidades de un pastizal del Norte de la Pampa Deprimida. XXII Reunión Argentina de Ecología.
Vecchio, M.C., Bolaños, V.A., Golluscio, R.A. y Rodríguez, A.M. (2019). Rotational grazing and exclosure improves grassland condition of the halophytic steppe in Flooding Pampa (Argentina) compared with continuous grazing. The Rangeland Journal, 41, 1-12.
Weatherspark. (2016a). El clima en Magdalena. https://es.weatherspark.com
Weatherspark. (2016b). El clima en Verónica. https://es.weatherspark.com
Wilson, A.M., Hyder, D.N. y Briske, D.D. (1976). Drought resistance characteristics of blue grama seedlings. Agronomy Journal, 68, 479-484.

