Susceptibilidad al daño por heladas en el pino híbrido, Pinus elliottii Engelm var. elliottii × Pinus caribaea Morlet var. hondurensis (sénéclauze) W.H. Barrett & Golfari y su relación con variables de crecimiento y fenología
Revista de la Facultad de Agronomía
Universidad Nacional de La Plata, Argentina
ISSN: 1669-9513
Periodicidad: Frecuencia continua
vol. 122, e132, 2023
Recepción: 10 Febrero 2023
Aprobación: 29 Mayo 2023
Resumen: Las especies híbridas se adaptan mejor a diferentes ambientes que sus progenitores. El pino híbrido (pino híbrido PEE×PCH) entre Pinus elliottii Engelm var. elliottii × Pinus caribaea Morlet var. hondurensis (Sénéclauze) W.H. Barrett & Golfari ha mostrado buen desempeño en regiones subtropicales de Australia, Brasil, Estados Unidos, Sudáfrica y Argentina. Sin embargo, su principal limitante en la expansión del área de plantación es la susceptibilidad del material al daño generado por las bajas temperaturas. Para ello se evaluó por medio de un análisis de componentes principales, como los caracteres asociados a crecimiento, desarrollo fenológico y diferentes condiciones de micrositio afectan a 3 familias de pino híbrido PEE×PCH y a su parental femenino Pinus elliottii. Se encontró un efecto diferencial del daño por bajas temperaturas a nivel sitio de plantación (BAJO y ALTO), especie (PEE y PEE×PCH), pero no a nivel de familia híbrida (F12, F28 y F51) afectando a todas por igual. Su crecimiento inicial y la rápida respuesta a condiciones favorables de crecimiento, son los principales factores que causan la susceptibilidad a bajas temperaturas.
Palabras clave: heladas, bajas temperaturas, mejora genética, fenología, ACP.
Abstract: The hybrid pine (hybrid pine PEE×PCH) between Pinus elliottii Engelm var. elliottii × Pinus caribaea Morlet var. hondurensis (Sénéclauze) W.H. Barrett & Golfari has performed well in subtropical regions of Australia, Brazil, United States, South Africa and Argentina. However, its main limitation in the expansion of the planting area is the susceptibility of the material to damage generated by low temperatures. For this purpose, it was evaluated by means of a principal component analysis, how the characters of growth, phenological development and different microsite conditions affect 3 hybrid families PEE×PCH and their female parent Pinus elliottii. A differential involvement of the damage by low temperatures was found at the plantation site level (LOW and HIGH), at the species level (PEE and PEE×PCH), but not at the hybrid family level (F12, F28 and F51) affecting all similarly. Their initial growth and rapid response to favorable growth conditions are the main factors that cause the susceptibility to low temperatures.
Keywords: frost, low temperatures, genetic improvement, phenology, PCA.
Introducción
Las bajas temperaturas determinan en gran medida la distribución natural de las especies vegetales (Poirier et al., 2010; Charrier et al., 2015; Turchetto et al., 2020), influyendo en el establecimiento, crecimiento y desarrollo de los cultivos (Inouye, 2000; Kodad y Socias i Company, 2005; Arias et al., 2015). Estas manifiestan diferentes respuestas adaptativas que les permiten sobrevivir a las bajas temperaturas (Thomashow, 1999) como también minimizar el daño generado por las bajas temperaturas (Charrier et al., 2015).
Hacen frente a estos eventos de estrés térmico mediante una combinación de mecanismos (Arias et al., 2015), pudiendo estar relacionados con estrategias que buscan evitar o controlar parcialmente la formación de hielo en la estructura interna de las plantas (evitar o tolerar, respectivamente) (Chaar, 2013; Charrier et al., 2015). El proceso de aclimatación al frío se ha estudiado ampliamente en especies modelos como ser Arabidopsis thaliana como así también en especies de interés agronómico y forestal (Mayr et al., 2006; Man et al., 2009; Charrier et al., 2014; Oberschelp et al., 2020).
La producción de híbridos entre especies del género Pinus que promueven mayor crecimiento, características deseables de madera, como así también resistencia a estrés y enfermedades está siendo ampliamente investigada (Malan, 1995; Dungey, 2001; Cerda Granados, 2012; Filippetti et al., 2014; Mabaso et al., 2019). Un criterio relevante al momento de producir híbridos que incluyen especies tropicales es reducir el riesgo a que el material vegetal se vea afectado por eventos de estrés por bajas temperaturas.
El híbrido entre Pinus elliottii Engelm var. elliottii × Pinuscaribaea Morlet var. hondurensis (Sénéclauze) W.H. Barrett & Golfari (de ahora en más, híbrido PEE×PCH) ha mostrado un buen desempeño en regiones subtropicales de Australia (Slee, 1972), Brasil y Estados Unidos (Nikles, 2017), Argentina (Belaber et al., 2018), Sudáfrica (Nilsson et al., 2020) y en otros diferentes países donde se lo ha cultivado. En Argentina las primeras plantaciones de híbrido PEE×PCH se iniciaron a principio de la década del 90 (Schenone y Pezzutti, 2003). Actualmente, la superficie cultivada en el país supera las 21.000 hectáreas (Cappa et al., 2013; Belaber et al., 2018) las cuales están concentradas en las Provincias de Misiones y Corrientes. La principal limitante para la expansión del área de plantación es la susceptibilidad del material al daño generado por las bajas temperaturas (Mabaso et al., 2019).
Según el sexto informe del IPCC (2022), se prevé un aumento en la intensidad y frecuencia de eventos cálidos y una disminución de la intensidad y frecuencia de eventos fríos en la región sur-este de América del Sur. Para la región mesopotámica Argentina, zona de implantación del híbrido PEE×PCH, se observa un aumento en las temperaturas medias mensuales como así también temperaturas mínimas absolutas superiores a las históricas (SMN, 2020). Si bien estos cambios generan condiciones favorables para el crecimiento de las plantas, también las expone a riesgos de daños por heladas (Gunderson et al., 2012; Pardos et al., 2014). Un desajuste entre el patrón de crecimiento y el clima, puede dar como resultado, menores tasas de crecimiento o provocar un daño grave si se produce un crecimiento activo en períodos desfavorables (Slee, 1972). Estas condiciones cambiantes del clima a escala regional podrían generar en el híbrido PEE×PCH un incremento en el riesgo de exposiciones a temperaturas que afecten su normal desarrollo, sumando un nuevo desafío al ya planteado anteriormente relacionado con la expansión de su área de implantación.
El objetivo de este trabajo es determinar si existe una estrategia de escape como mecanismo de aclimatación a las bajas temperaturas en familias de pino híbrido PEE×PCH, y determinar si las condiciones climáticas asociadas a las características del micrositio influyen en el riesgo de afectación por heladas en el primer año de plantación.
Metodología
material vegetal y SITIO de estudio
El material vegetal utilizado consistió en tres bulk de plantines obtenidos de estacas enraizadas (cuttings) de 13 meses de edad de pino híbrido PEE×PCH, provenientes de 3 familias (F12, F51, F28) y plantines obtenidos de un bulk de semillas (seedling) de Pinus elliottii var. elliottii (PEE). Ambos materiales fueron brindados por el Programa de Mejoramiento Genético Forestal (PMGF) de la Estación Experimental Agropecuaria Montecarlo del Instituto Nacional de Tecnología Agropecuaria (INTA). Las 3 familias de híbrido F. de PEE×PCH utilizadas en este estudio se obtuvieron de cruzamientos controlados realizados en el huerto semillero clonal de PEE ubicado en el Campo Anexo Manuel Belgrano (CAMB) del INTA, en la Localidad de San Antonio, Misiones. Este material híbrido F. PEE×PCH INTA-PINDO es producto de un convenio entre el INTA y la empresa PINDO S.A.
El clima según la clasificación de Köppen es de tipo Cfa, templado húmedo sin estación seca, macro térmico, constantemente húmedo y subtropical con ocurrencia de heladas (Boca et al., 2017). La precipitación anual promedio es de 1865,4 ± 456,4 mm periodo 1927-2018 (Silva, et al., 2019a). El suelo corresponde a un rojo profundo arcilloso, bien drenado y fuertemente ácido (pH 6,06), representado por un complejo de Kandiudultes y Kandiudalfes ródicos (Von Wallis et al., 2020).
El día 2 de julio del 2020 se instalaron 2 ensayos en el Campo Anexo Laharrague (CAL) del INTA en la Localidad de Montecarlo, Misiones (26°33'31,57"S y 54°40'28,51"O). El lote presenta una pendiente del 3% en sentido norte-sur. Uno de los sitios de ensayo se instaló en la parte alta (ALTO) del terreno, y el otro en la parte baja (BAJO) con una separación de 200 metros entre ellos con el objetivo de captar la variación de las condiciones climáticas asociados al micrositio que pueden afectar el normal desarrollo del híbrido PEE×PCH. Previo a la plantación el terreno fue desmalezado y subsolado a 0,50 metros de profundidad.
El diseño utilizado para la instalación de los ensayos consistió en cuatro bloques completos al azar, con cinco árboles por parcela, totalizando un número efectivo de 160 plantas entre ambos ensayos. El marco de plantación fue de 3 metros entre líneas por 1 metro entre plantas.
Variables evaluadas
Variables climáticas
En cada ensayo (ALTO y BAJO) se registró la temperatura ambiente (T. °C) y la humedad relativa del aire (H. %) mediante sensores HOBBO U23-001 (Pro series, Onset Computer Corporation, Pocasset, MA, USA) instalados a 0,30 metros del suelo. Las variables se registraron diariamente cada 30 minutos. Se determinó la amplitud térmica diaria (A., °C) como la diferencia entre las temperaturas máxima y mínima absolutas. Se obtuvieron los valores climáticos históricos del periodo 1965-2018. Para el periodo 1965-2011 los datos provenían del registro de termo-higrógrafo ubicado en abrigo meteorológico tipo A. Para el periodo 2012-2013 de la estación meteorológica automática marca Davis y para el periodo 2013-2018 de la estación automática marca Nimbus, todos ubicados en el CAL (Silva et al., 2019b). Se registraron los días con eventos de temperaturas inferiores a 0°C (heladas).
Fenología
El desarrollo de las plantas fue evaluado mediante la determinación del estado fenológico de la brotación de la yema terminal (Estado_feno). Para ello se elaboró una escala con cinco categorías (de 0 a 4) siendo 0 una yema cerrada y 4 una yema completamente abierta (Tabla S1, material suplementario). La confección de dicha escala se realizó por medio de observaciones de las plantas a campo en intervalos de 2 días. El periodo de observación fue entre julio y septiembre para los sitios ALTO Y BAJO.
Variables dasométricas
La altura total (AT, cm), el diámetro a la altura del cuello (DAC, mm) y la sobrevivencia (SOB, %) fueron evaluados en todas las plantas de ambos sitios. La AT con regla metálica, el DAC con calibre digital (ESSEX, 150 mm) y la SOB con una escala de 0 y 1 (donde 0 indica planta muerta y 1 planta viva), utilizando para el registro de cada variable una colectora de datos (SPECTRA NOMAD 900). Las variables dasométricas fueron categorizadas en estáticas y dinámicas. Las estáticas corresponden a la AT y DAC de cada planta al momento de la plantación (AT. y DAC.), y posterior a la ocurrencia de las heladas registradas en el invierno del año de plantación (AT55 y DAC55). Las variables dinámicas corresponden a los Incrementos parciales (Inc) en AT y DAC producido dentro del periodo de evolución de las variables estáticas (Arias et al., 2019). Se utilizaron para ello las variables estáticas; tomando como inicio la AT. y el DAC. y como fin la AT55 y el DAC55.
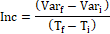
donde
Var. corresponde a AT55 y DAC55.
Var. corresponde a AT. y DAC..
T. número de días trascurridos hasta medición posterior a los eventos de heladas (día 55).
T. momento de instalación del ensayo (día 0).
Además de las variables estáticas y dinámicas se calculó la unidad de Esbeltez (E, cm mm-1) utilizando la metodología de (León-Sánchez et al., 2019). Esta variable se determinó para AT. y DAC. (E.) como también AT55 y DAC55 (E55).
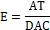
Afectación por heladas
La afectación por heladas fue visualmente evaluada en 2 categorías, 2 días posteriores a la ocurrencia de temperaturas por debajo de 0°C registrada en ambos sitios. Las plantas afectadas (plantas con pérdida de turgencia de las acículas que rodean la yema apical) fueron registradas con el valor 1 y las plantas no afectadas con el valor 0 (Figura S2, material suplementario).
Análisis de datos
Un Análisis de Componentes Principales (ACP) fue realizado para identificar las variables que explican la posible agrupación por similitud (Batalla, 2019). Las variables que se incluyeron en un primer análisis fueron AT., AT55, DAC., DAC55, E., E55, Inc_AT e Inc_DAC. Se determinó una alta correlación entre AT. y AT55; DAC.y DAC55 y E. y E55. Siendo que las variables iniciales (AT., DAC. y E.) maximizan sobre las dos primeras dimensiones del ACP el porcentaje de variación explicado por el conjunto de variables estudiadas, solo las variables que describen el estado inicial al momento de plantación fueron usadas para el ACP. Sitio, afectación, especie y familia fueron usados como variables cualitativas suplementarias con el fin de establecer los agrupamientos por similitud. Se realizó un análisis de varianza para establecer la existencia de diferencias estadísticas significativas. Se consideraron los factores Sitio, Especies, Bloques anidados dentro de Sitio y Familia anidado dentro de Especies. La unidad experimental fue la parcela. La trasformación Log10 fue aplicada a los incrementos de AT y DAC previo a su análisis. Todos los análisis estadísticos se realizaron en el entorno R (R Core Team 2020), mediante el uso de los siguientes paquetes estadísticos y herramientas gráficas (FactoMineR; factoextra, vcd, ggplot2).
Resultados
Temperaturas aMbientales diarias
En la Tabla 1 se presentan los datos climáticos registrados en los sitios ALTO, BAJO e históricos (1965-2018) recolectados de la casilla meteorológica del Campo Anexo Laharrague, ubicado a 800 metros del sitio de plantación.
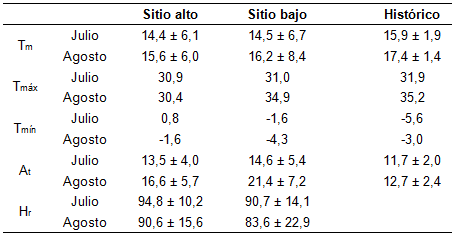
TABLA 1
Registro de variables ambientales en los sitios ALTO y BAJO en julio y agosto del 2020 y datos históricos de una serie de 46 años. Tm: temperatura media, At: amplitud térmica y Hr: humedad relativa. Valores promedio ± desvíos estándar. Tmáx: temperaturas máximas absoluta y Tmín: temperaturas mínimas absolutas.
La temperatura media para los meses de julio y agosto en los sitios ALTO y BAJO fueron similares entre sí (Tabla 1), y a su vez similares a los valores medios históricos. Los valores promedios diarios mostraron la misma tendencia (Figura 1C), registrándose valores muy similares en ambos sitios.
Las temperaturas máximas absolutas de los sitios de plantación durante el mes de julio fueron similares entre ellos y con la histórica registrada. Durante el mes de agosto, en el sitio BAJO se registró una máxima absoluta superior en 4,5 °C muy próxima a los valores históricos registrados (Tabla 1). Este evento de temperaturas máximas superiores tuvo una duración de 14 días (Figura 1B), presentándose en ambos sitios, siendo en el sitio BAJO más extremo.
Los datos históricos exhiben temperaturas por debajo de los 0°C para los meses de julio y agosto (-5,6 ° C y -3,0 °C, respectivamente). En el mes de julio, tanto en el sitio ALTO como en el BAJO, los valores mínimos registrados indican un mes más benévolo comparado con los datos históricos, sin heladas registradas en el sitio ALTO. Sin embargo, en el sitio BAJO (sitio dónde se registraron signos de afectación de brotes terminales en plantas), se registró en agosto una mínima de -4,3 °C, mientras que en el ALTO fue de -1,6 °C (Tabla 1).
La amplitud térmica del mes de julio en los sitios de plantación fue similar a los valores históricos reportados. En el mes de agosto se registró un aumento en esta variable (16,6 ± 5,7 y 21,4 ± 7,2, sitio ALTO y BAJO, respectivamente) en relación a los valores históricos registrados (Tabla 1). Este fenómeno se registró en ambos sitios, siendo más alto en el sitio BAJO (Figura 1D). La humedad relativa (registrada solamente en los sitios de plantación) fue alta, siendo siempre mayor en el sitio ALTO.
En relación a los eventos de bajas temperaturas, en el sitio ALTO se registraron en total 7 días con temperaturas menores o iguales a 2,5°C (heladas agrícolas), (Silva et al., 2019a), de los cuales, 4 días fueron en el mes de julio (días 3, 15, 29 y 30) y 3 en el mes de agosto (días 20, 21 y 22). Solamente el día 21 se registró temperatura inferior a los 0°C (heladas). En el sitio BAJO se registraron 10 días con temperaturas iguales o inferiores a 2,5°C, distribuidos 6 días en el mes de julio (3, 4, 10, 15, 29 y 30) y 4 días en agosto (días 20, 21, 22 y 23). Registrándose el día 30 de julio el primer día de temperaturas inferiores a 0°C (heladas), seguidos de los días 20, 21 y 22 de agosto con heladas próximas a los valores históricos, tal cual se indicó precedentemente (Figura 1A).
En la Figura 1 se muestran los valores diarios de las temperaturas registradas en los sitios ALTO y BAJO durante los meses de julio y agosto de 2020.
estado fenológico
El híbrido PEE×PCH no evidenció un estado de latencia como sí se determinó en PEE. El estado de la yema terminal del híbrido PEE×PCH cambió de una condición de latencia (categoría 0-1), a un estado de plenitud de desarrollo (categoría 3-4) en solamente 50 días. Por el contrario, en PEE la latencia fue más notoria, pasando de una categoría 0-1 a 1-2 para el mismo periodo de tiempo (Figura 2A, 2B y 2C).
Crecimiento y afectación por bajas temperaturas
La sobrevivencia (SOB) fue del 100% en ambos sitios evaluados.
En el sitio BAJO, el 20% de las plantas afectadas por bajas temperaturas corresponden solamente al híbrido PEE×PCH, mientras que en el sitio ALTO no se encontraron plantas afectadas.
Los caracteres de DAC y AT (iniciales y finales, como así los incrementos parciales) y el estado fenológico explican el 67,1 % de la variabilidad total en las dos primeras dimensiones (Dim1 y Dim2) del ACP (ambos sitios). Los caracteres que más contribuyen a la explicación de la variabilidad en la Dim1 son E. (33,8%), Inc_AT (23,6%) (correlacionados negativamente entre ellos) y AT. (21,6%). Mientras que en la Dim2 son DAC. (29,9%), Estado_feno (24,0%) y AT. (23,1%). Las elipses de confianza para los sitios ALTO y BAJO usados como factores cualitativos complementarios de clasificación se superponen (Figura 3).
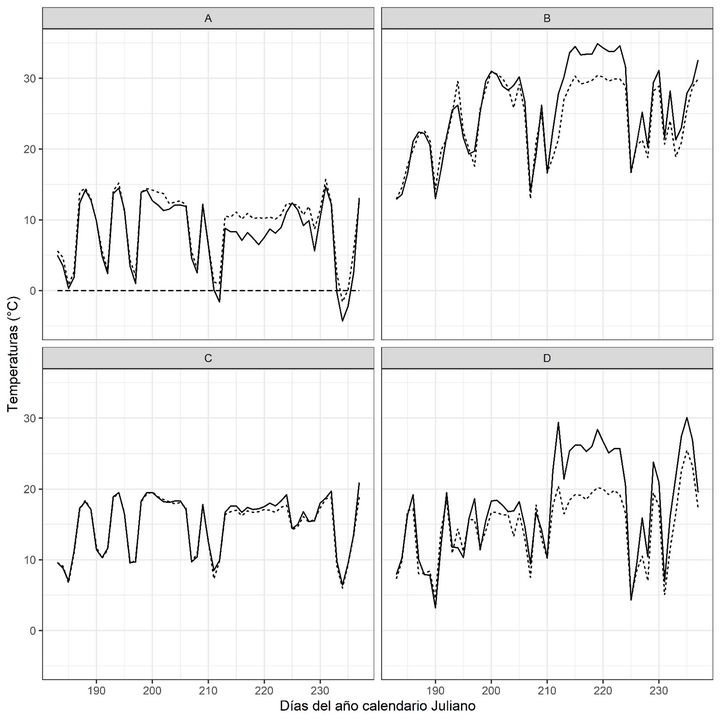
FIGURA 1
(A) Temperaturas mínimas absolutas (se indica línea de 0°C), (B) máximas absolutas, (C) medias y (D) amplitud térmica de los sitios de plantación. Líneas continuas: sitio BAJO, líneas discontinuas: sitio ALTO.
En el sitio BAJO, las 2 primeras dimensiones del ACP explican el 70,3% de la variabilidad total. La afectación por bajas temperaturas (factor cualitativo complementario usado como criterio de clasificación), discriminan las plantas afectadas de las no afectadas (Figura 4A) según su AT., DAC. y Estado_feno. El DAC. se relaciona positivamente con Estado_feno (fenología de crecimiento del brote). Mientras que los incrementos en altura y diámetro (Inc_AT e Inc_DAC, respectivamente) se asocian negativamente con DAC. y ATi, siendo las variables que contribuyen al agrupamiento por similitud de las plantas sin signos de afectación por heladas.
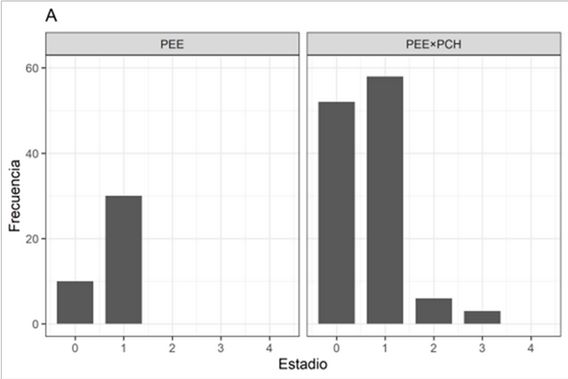
FIGURA 2
Evolución de desarrollo de la fenología del brote apical del híbrido PEE×PCH y su parental femenino PEE. Estado fenológico a los 11 (A), 27 (B) y 50 (C) días de instalación de los ensayos.
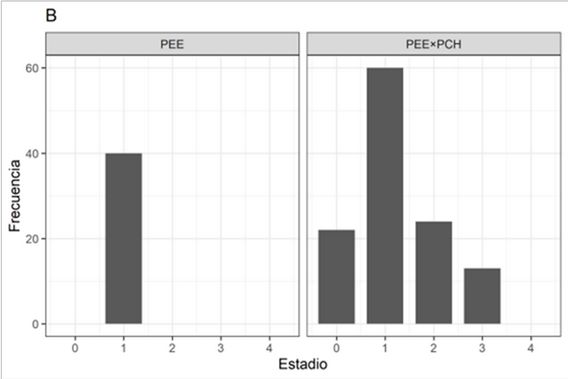
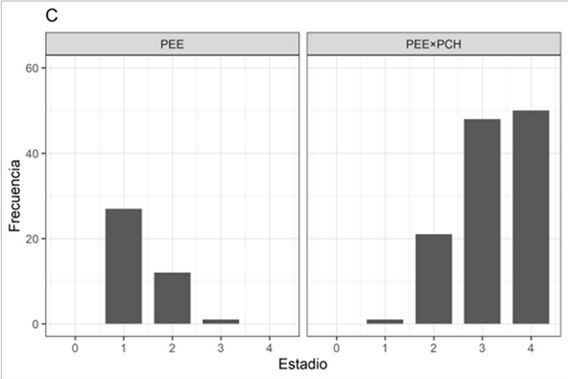
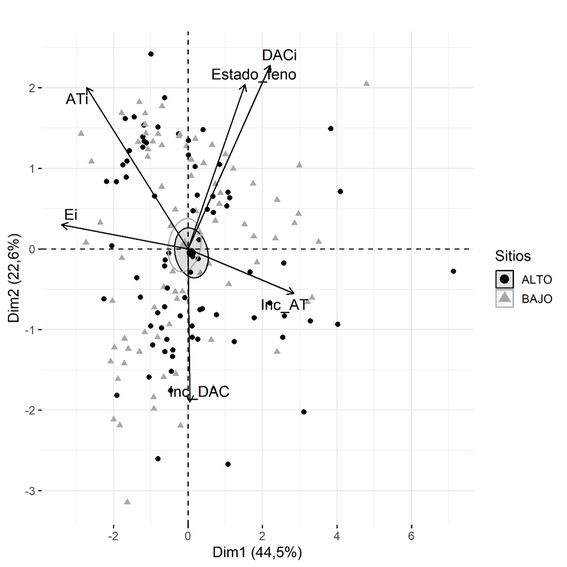
FIGURA 3
Correlaciones de los componentes principales de las variables de crecimiento y fenología en ambos sitios, agrupamiento individual y elipses de confianza a nivel sitio.
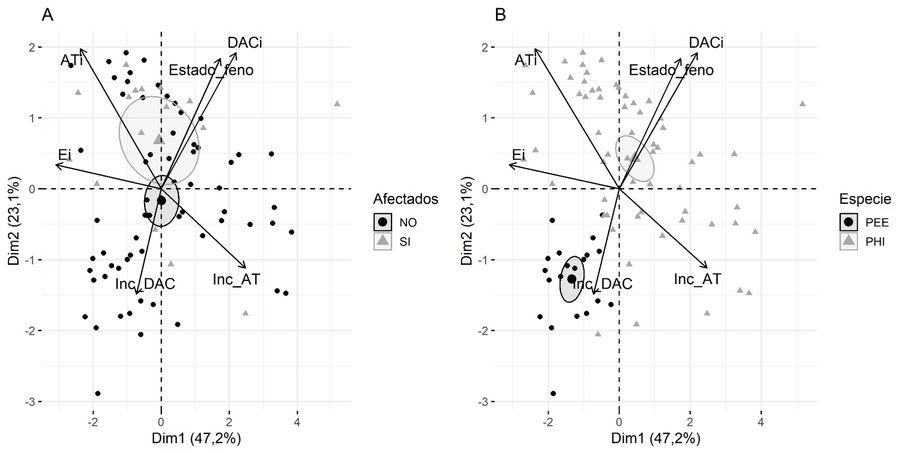
FIGURA 4
Correlaciones de los componentes principales de las variables de crecimiento en el sitio BAJO. Porcentajes de variabilidad que se logra explicar en cada dimensión. Criterios de clasificación: (A) Afectación. (B) Especies.
Al utilizar Especie como factor de clasificación, las elipses de confianza discriminan con mayor precisión a las plantas, siendo la Especie híbrido PEE×PCH asociada con las variables AT., DAC. y Estado_feno (Figura 4B), variables que a la vez se asocian con plantas afectadas por las bajas temperaturas.
En la Figura 5 se observa como las elipses de confianza para las distintas familias se asocian a diferentes variables. La familia F12 (híbrido PEE×PCH) se asocia a la variable AT., mientras que la familia F51 (híbrido PEE×PCH) muestra una asociación con el Inc_AT. El material parental PEE se asocia al Inc_DAC.
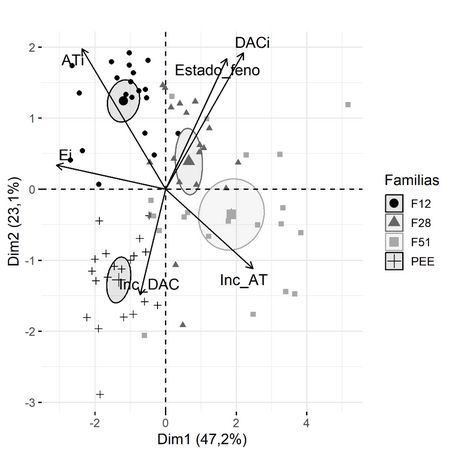
FIGURA 5
Correlaciones de los componentes principales de las variables de crecimiento en el sitio BAJO. Porcentajes de variabilidad que se logra explicar en cada dimensión. Criterios de clasificación: Familias.
ANOVA
Un análisis de varianza realizado para cada una de las variables dasométricas aquí representadas en el ACP, demuestra efectos significativos entre Especie para DAC., DAC55 y log10(Inc_AT). El efecto del bloque (anidado en Sitio) fue significativo para AT., AT55, DAC55 y el log10(Inc_AT), mientras que la Familia (anidada en Especie) fue significativa para todas las variables, excepto para DAC. y DAC55. Sin embargo, no se establecieron diferencias significativas para ninguna de las variables a nivel Sitio (Tabla S2, material suplementario). La Parcela fue considerada como unidad experimental, con lo cual la estimación de los cuadrados medios de Parcela fueron los utilizados para estimar los valores de . y su probabilidad asociada. De esta manera evitamos de correr el riesgo de rechazar la hipótesis nula cuando esta es en realidad verdadera (Error de tipo I o error de la unidad experimental).
Discusión
Relación entre el micrositio y la fenología de brotación
La variabilidad climática asociada a la frecuencia e intensidad de heladas fue capturada entre los sitios de plantación, separados solamente 200 metros uno del otro. Los registros climáticos indican un comportamiento diferencial en relación a los valores absolutos mínimos de temperatura (heladas más intensas), a la vez que temperaturas máximas más altas, generando condiciones de clima con mayor amplitud térmica para el sitio BAJO. Es en este sitio dónde se observaron signos de afectación por bajas temperaturas.
Por tratarse de una superficie cóncava, las condiciones de relieve favorecen la acumulación del aire frío por la noche (Marquis et al., 2021). Por el contrario, durante el día su exposición al sol por más tiempo (ya que no era cubierto por la sombra de las plantaciones lindantes como si el sitio ALTO) generó un aumento en las temperaturas máximas. Entre los dos eventos de temperaturas por debajo de los 0°C, el sitio BAJO presentó 14 días con amplitudes térmicas superiores al sitio ALTO. Según el reporte del clima del Servicio Meteorológico Nacional (SMN, 2020) a nivel país se observó una tendencia similar durante el invierno, donde predominaron mayores amplitudes térmicas. Esto fue el resultado de temperaturas máximas absolutas más elevadas y mínimas absolutas inferiores en ese periodo.
Las condiciones de microclima, como ser mayores amplitudes térmicas y temperaturas mínimas absolutas más bajas, afectaron el híbrido PEE×PCH, donde se evidenciaron signos de afectación por bajas temperaturas. Esto podría estar influenciado por su capacidad de responder de manera inmediata a condiciones favorables para su crecimiento, las cuales se registraron entre los 2 eventos de heladas. Según Turchetto et al. (2020), la combinación de un crecimiento continuo y cambios bruscos de temperatura incrementa los riesgos de afectación frente a eventos de bajas temperaturas, más aún en sitios donde las mínimas absolutas rondan los -5°C.
El escape a las bajas temperaturas está relacionado al estado fenológico de los brotes terminales de las plantas. En el híbrido PEE×PCH, el crecimiento del brote terminal no cesó, contrariamente a lo que si sucede en coníferas de clima templado (Viveros-Viveros et al., 2007, De Jesus-Reyes et al., 2020). Slee (1972), encontró que Pinuselliottii presenta durante el invierno una yema en evidente estado de reposo, mientras que el Pinuscaribaea y el híbrido PEE×PCH, presentaron una yema blanquecina y suculenta, lista para desarrollarse. Igual comportamiento fue observado en este trabajo, donde P. elliottii manifestó un mecanismo de escape a bajas temperaturas. Las coníferas pueden adquirir mayor grado de resistencia al frío con días cortos y temperaturas ambiente templadas, donde las bajas temperaturas aceleran la adquisición de resistencia al frío (Lu et al., 2007).
Al igual que lo reportado en este trabajo, Marquis et al. (2021) demostraron que los daños por heladas en plantas están asociado a las condiciones del micrositio en la que se encuentran instaladas. La acumulación de aire frío en los sitios bajos, si bien retrasan el crecimiento, también favorecen a generar resistencia a las bajas temperaturas. Por el contrario, el incremento de las temperaturas (máximas absolutas y heladas más intensas y frecuentes) afectan negativamente en el híbrido PEE×PCH su capacidad de resistencia a las bajas temperaturas.
Las variables dasométricas y su relación frente a las bajas temperaturas
El análisis de ACP permitió corroborar que las variables dasométricas no pueden ser discriminadas al utilizar el factor sitio como criterio de clasificación. Sus elipses de confianza se superponen. Ambas cubren un área similar, demostrando así que el conjunto de variables estáticas, dinámicas y fenológicas analizadas en este trabajo son similares en ambos sitios. Sin embargo, al utilizar a la afectación como variable de clasificación, se puede observar una clara separación entre las elipses de confianza; trasladándose la elipse correspondiente a las plantas afectadas hacia valores positivos de la Dim2 del ACP realizado para el sitio BAJO. Esto indica una asociación positiva con las variables DAC.; Estado_feno y AT. las cuales son las que explican la mayor variabilidad en esa dimensión.
Las 3 familias ensayadas en este trabajo mostraron signos de afectación por bajas temperaturas, mientras que su parental femenino (PEE) no. El mayor número de individuos afectados de la familia F12 (40%) en el sitio BAJO, se relacionó con la mayor altura que presentaron al momento de ocurrencia de las heladas. Se sabe que las condiciones de la superficie del suelo, al momento de producirse las heladas, determinan un micro clima y es donde se dan los mayores gradientes de temperatura (Enz & Hofman, 2019). Las temperaturas nocturnas de un suelo descubierto son superiores a las de un suelo cubierto. Es por ello que las plantas que superen en altura este microclima generado en los primeros centímetros del suelo, quedarán más expuestas a las bajas temperaturas en el primer año de plantación. En este sentido, hemos comprobado que la familia F51 del híbrido PEE×PCH, de menor crecimiento fue la menos afectada por las bajas temperaturas, con solamente el 15 % de las plantas del sitio BAJO.
En el caso del parental femenino PEE, de menor crecimiento, menor incremento en altura y con latencia vegetativa, combinaciones que favorecen la mayor resistencia al frío, tiene un comportamiento similar al reportado por De Jesus-Reyes et al. (2020) en Pinus radiata D. Don., donde las poblaciones con menores tasas de crecimiento absolutas mostraron una mayor resistencia. Viveros-Viveros et al., (2007) encontraron un comportamiento similar para varias especies del género Pinus.
En el análisis de ACP en el sitio BAJO, el criterio de clasificación Especie discrimina el híbrido PEE×PCH claramente del material parental PEE. A su vez, las familias híbridas PEE×PCH manifiestan una asociación positiva con caracteres relacionados al crecimiento (AT., DAC., Estado_feno e Inc_AT). Mientras que, el parental PEE, expresa una asociación negativa con estas variables, pero positiva con el Inc_DAC. La familia híbrida PEE×PCH F28 y el parental PEE presentan alturas similares. Sin embargo, el 25 % de las plantas de la familia F28 fueron afectadas por las bajas temperaturas. El crecimiento vegetativo continuo del híbrido PEE×PCH lo expone a condiciones en las cuales las bajas temperaturas afectan las yemas terminales. La no cesación del crecimiento explica las mayores tasas de crecimiento. Slee (1972) encontró que el híbrido PEE×PCH presenta un patrón de crecimiento intermedio a sus progenitores, donde el Pinus elliottii al llegar el invierno entra en latencia, mientras que el Pinus caribaea solamente disminuye su tasa de crecimiento.
Conclusiones
La rápida respuesta del pino híbrido PEE×PCH frente a condiciones favorables de crecimiento se refleja a través de su fenología. No se identificó un cese del crecimiento. Esto se relaciona negativamente con la resistencia a bajas temperaturas.
A pesar que la temperatura mínima absoluta registrada de -4,6 °C no provocó mortandad de plantas, tampoco daños suficientes que afecten el normal desarrollo de las mismas, las diferentes temperaturas mínimas absolutas registradas en los sitios de estudios indican que la correcta selección del sitio es el criterio principal a considerar al momento de la plantación del híbrido PEE×PCH.
Además, la existencia de una amplia base genética en el programa del híbrido F. PEE×PCH INTA-PINDO, conformada por más de 400 familias y su potencial evaluación ecofisiológica permitirían identificar materiales que se adapten mejor a las bajas temperaturas.
La fenología del híbrido PEE×PCH no demuestra una estrategia de escape a las heladas, por lo que su resistencia a las bajas temperaturas está mayormente asociada a caracteres fisiológicos de tolerancia. La correcta elección del sitio de plantación es una opción para reducir el daño que generan las bajas temperaturas.
Agradecimientos
Los autores agradecen al Instituto Nacional de Tecnología Agropecuaria Estación Experimental Agropecuaria Montecarlo y a la empresa PINDO S.A. quienes pusieron a disposición el material vegetal y el apoyo logístico para establecer y medir estos ensayos. Agradecemos a la Ingeniera Forestal Iris Figueredo y al Sr. Diego Bogado por la colaboración en el trabajo de campo.
Material suplementario
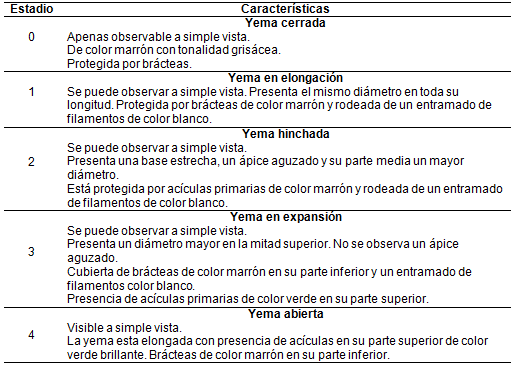
TABLA S1
Descripción de la escala de los estadios fenológicos en la etapa de brotación utilizada para la evaluación del híbrido PEE×PCH.

FIGURA S1
Efecto de las heladas sobre plantas de PEE×PCH en el sitio BAJO. (A) perdida de turgencia en acículas y brote apical, corresponde a categoría 1 en la escala de evaluación de la afectación. (B) planta no afectada por las heladas, corresponde al valor 0 en la escala de evaluación de la afectación.
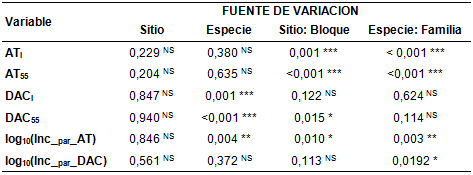
TABLA S2
Valor de probabilidad y nivel de significancia para las variables dasométricas ATi: altura total inicial, AT55: altura total en el día 55, DACi: diámetro a la altura del cuello inicial, DAC55: diámetro a la altura del cuello en el día 55, log10(Inc_AT): logaritmo en base 10 del incremento parcial en altura total, log10(Inc_DAC): logaritmo en base 10 del incremento parcial en diámetro a la altura del cuello, considerando como fuentes de variación el sitio, la especie, el bloque dentro del sitio y la familia dentro de la especie. La Parcela fue considerada unidad experimental. La estimación de los cuadrados medios de parcela fueron los utilizados para estimar los valores de F y su probabilidad asociada. Códigos de significancias: <0,001 ‘***’ 0,001 ‘**’ 0,01 ‘*’ no significativo ‘NS’
FIGURA S2
Frecuencias absolutas de distribución de las variables dasométricas estáticas (A) ATi, (B) AT55, (C) DACi y (D) DAC55
FIGURA S3
Frecuencias absolutas de las variables dasométricas dinámicas. (A) log10(Inc_AT) y (B) log10(Inc_DAC).
Bibliografía
Arias, R.C., Reyes, J.J., Ray, J.V., Benítez, D.G., Hernández, L.G., y Ledea, J.L. (2019). Indicadores morfométricos en nuevas variedades megatérmicas de Cenchrus purpureus tolerantes al estrés hídrico. Tropical and Subtropical Agroecosystems, 22, 115–125.
Arias, N.S., Bucci, S.J., Scholz, F.G., y Goldstein, G. (2015). Freezing avoidance by supercooling in Olea europaea cultivars: The role of apoplastic water, solute content and cell wall rigidity. Plant, Cell and Environment, 38(10), 2061–2070. https://doi.org/10.1111/pce.12529.
Batalla, G.A. (2019). Evaluación de la fenología de plantas cultivadas de Yerba Mate (Ilex paraguariensis St. Hil) como base para un modelo predictivo de rendimiento de hoja verde. Tesis de Maestría, Facultad de Ciencias Agrarias. Universidad Nacional del Nordeste.
Belaber, E.C., Gauchat, M.E., Reis, H.D., Borralho, N.M., y Cappa, E.P. (2018). Genetic Parameters for Growth, Stem Straightness, and Branch Quality for Pinus elliottii var. elliottii × Pinus caribaea var. hondurensis F 1 Hybrid in Argentina. Forest Science, 64(6), 595–608. https://doi.org/10.1093/forsci/fxy021.
Boca, T., Fassola, H., Crechi, E., Barth, S., Keller, A., Winck, R., y Ferrere, P. (2017). Modelos aditivos de predicción de biomasa aérea de Pinus elliottii var. elliottii × Pinuscaribaea var. hondurensis de Misiones Argentina. Quebracho, 25 (1,2), 5–15.
Cappa, E.P., Marcó, M., Garth Nikles, D., y Last, I.S. (2013). Performance of Pinus elliottii, Pinus caribaea, their F., F. and backcross hybrids and Pinus taeda to 10 years in the Mesopotamia region, Argentina. New Forests, 44(2), 197–218. https://doi.org/10.1007/s11056-012-9311-2.
Cerda Granados, A.D. (2012). Geographical Variation of Cold Hardiness in Pinus patula Provenances and Genetic Inheritance of Cold Hardiness in Pinus patula × Pinus tecunumanii Hybrids. Thesis of Master, Faculty of North Carolina State University.
Chaar, J.E. (2013). Resistencia a heladas en plantas frutales. Avances en Investigación Agropecuaria, 17(3), 109-121.
Charrier, G., Charra-Vaskou, K., Kasuga, J., Cochard, H., Mayr, S., y Ameglio, T. (2014). Freeze-Thaw Stress: Effects of Temperature on Hydraulic Conductivity and Ultrasonic Activity in Ten Woody Angiosperms. Plant Physiology, 164(2), 992-998. https://doi.org/10.1104/pp.113.228403.
Charrier, G., Ngao, J., Saudreau, M., y Améglio, T. (2015). Effects of environmental factors and management practices on microclimate, winter physiology, and frost resistance in trees. Frontiers in Plant Science, 6, 1-18. https://doi.org/10.3389/fpls.2015.00259.
De Jesus-Reyes, G., Jesús Vargas-Hernández, J., Cruz-Huerta, N., y López-Upton, J. (2020). Populations of Pinus radiata D. Don differ in low-temperature tolerance and growth rate. Revista Chapingo, Serie Ciencias Forestales y Del Ambiente, 26(3), 469–484. https://doi.org/10.5154/r.rchscfa.2019.12.096.
Dungey, H.S. (2001). Pine hybrids a review of their use performance and genetics. Forest Ecology and Management, 148, 243-258. https://doi.org/10.1016/S0378-1127(00)00539-9.
Enz, J.W., Hofman, V. y Thostenson, A. (2019). Air temperature inversions. NDSU Extension, 1705, 1–16.
Filippetti, M., Guillaumet, A., y Manavella, R. (17, 18 y 19 de septiembre de 2014). Aptitud del hibrido Pinus elliottii var. elliottii × Pinus Caribaea var. hondurensis para la fabricación de vigas de madera laminada encolada. XXIII Jornadas Argentinas de Ingeniería Estructural. (pp.1-14). Buenos Aires.
Gunderson, C.A., Edwards, N.T., Walker, A.V., O’Hara, K.H., Campion, C.M., y Hanson, P.J. (2012). Forest phenology and a warmer climate growing season extension in relation to climatic provenance. Global Change Biology, 18(6), 2008–2025. https://doi.org/10.1111/j.1365-2486.2011.02632.x.
Inouye, D.W. (2000). The ecological and evolutionary significance of frost in the context of climate change. Ecology Letters, 3(5), 457-463. https://doi.org/10.1046/j.1461-0248.2000.00165.x.
IPCC (2022). IPCC Sixth Assessment Report. Recuperado el 15 de mayo de 2022 de: https://www.ipcc.ch/report/ar6/wg2/.
Kodad, O., y Socias i Company, R. (2005). Daños diferenciales por heladas en flores y frutos y criterios de selección para la tolerancia a heladas en el almendro. Información Técnica Económica Agraria, 101(4), 349–363.
León-Sánchez, M.A., Pozo, J.L. R., León, V.E.P., Vichot, M.B., y Herrero-Echavarría, G. (2019). Slenderness and mineral fertilization of Pinus caribaea plantations in Cuba. Madera y Bosques, 25(2), 1–11. https://doi.org/10.21829/myb.2019.2521777.
Lu, P., Colombo, S.J., y Sinclair, R.W. (2007). Cold hardiness of interspecific hybrids between Pinus strobus and P. wallichiana measured by post-freezing needle electrolyte leakage. Tree Physiology, 27(2), 243-250. https://doi.org/10.1093/treephys/27.2.243.
Mabaso, F., Ham, H., y Nel, A. (2019). Frost tolerance of various Pinus pure species and hybrids. Southern Forests, 81(3), 273-280. https://doi.org/10.2989/20702620.2019.1615212.
Malan, F.S. (1995). The basic wood properties and sawtimber quality of South African grown Pinus elliottii × Pinus caribaea. South African Forestry Journal, 173(1), 35–41. https://doi.org/10.1080/00382167.1995.9629689.
Man, R., Kayahara, G.J., Dang, Q.L., y Rice, J.A. (2009). A case of severe frost damage prior to budbreak in young conifers in Northeastern Ontario: Consequence of climate change? Forestry Chronicle, 85(3), 453–462. https://doi.org/10.5558/tfc85453-3
Marquis, B., Duval, P., Bergeron, Y., Simard, M., Thiffault, N., y Tremblay, F. (2021). Height growth stagnation of planted spruce in boreal mixed woods: Importance of landscape, microsite, and growing-season frosts. Forest Ecology and Management, 479, 1-12. https://doi.org/10.1016/j.foreco.2020.118533.
Mayr, S., Hacke, U., Schmid, P., Schwienbacher, F., y Gruber, A. (2006). Frost drought in conifers at the alpine timberline: Xylem dysfunction and adaptations. Ecology, 87(12), 3175-3185. https://doi.org/10.1890/0012-9658(2006)87[3175:FDICAT]2.0.CO;2
Nikles, D. (2017). Southern pines. Woods Wiki Short Stories. Compiled by Brown, AG. 1-5.
Nilsson, O., Hodge, G.R., Frampton, L.J., Dvorak, W.S., y Bergh, J. (2020). Growth and modulus of elasticity of pine species and hybrids three years after planting in South Africa. Southern Forests, 82(4), 367–376. https://doi.org/10.2989/20702620.2020.1733769.
Oberschelp, G.P.J., Guarnaschelli, A.B., Teson, N., Harrand, L., Podestá, F.E., y Margarit, E. (2020). Cold acclimation and freezing tolerance in three Eucalyptus species: A metabolomic and proteomic approach. Plant Physiology and Biochemistry, 154, 316–327. https://doi.org/10.1016/j.plaphy.2020.05.026.
Pardos, M., Climent, J., Almeida, H., y Calama, R. (2014). The role of developmental stage in frost tolerance of Pinus pinea L. seedlings and saplings. Annals of Forest Science, 71(5), 551–562. https://doi.org/10.1007/s13595-014-0361-9.
Poirier, M., Lacointe, A., y Améglio, T. (2010). A semi-physiological model of cold hardening and dehardening in walnut stem. Tree Physiology, 30(12), 1555-1569. https://doi.org/10.1093/treephys/tpq087.
Schenone, R.A., y R.V. Pezzutti. (2003). Productividad de progenies de Pinus elliottii × Pinuscaribaea var. hondurensis. Proc. of 12º Congreso Forestal Mundial. Québec, Canadá.
Silva, F., Haberle, T., y Muller, H. (2019a). Agroclimatología de heladas para la región de Montecarlo. XIII Jornadas Técnicas Forestales y Ambientales, (pp. 529–531). El Dorado, Misiones.
Silva, F., Haberle, T., Muller, H., Münz, R., y Visentini, J. (2019b). Normales del clima de la región de Montecarlo. XVIII Jornadas Técnicas Forestales y Ambientales. El Dorado, Misiones.
Slee, M.U. (1972). Growth caribbean patterns of slash and pine and their hybrids. Euphytica, 21(1), 129–142.
SMN. (2020). Estado del clima en la Argentina 2020. http://hdl.handle.net/20.500.12160/1559.
Thomashow, M.F. (1999). Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annual Review of Plant Physiology and Plant Molecular Biology, 50 (1), 571–599. https://doi.org/10.1146/annurev.arplant.50.1.571.
Turchetto, F., Araujo, M.M., Griebeler, A.M., Rorato, D.G., Pasquetti Berghetti, Á.L., Barbosa, F.M., y Santos de Lima, M. (2020). Can intensive silvicultural management minimize the effects of frost on restoration plantations in subtropical regions? Journal of Environmental Management, 269, doi https://doi.org/10.1016/j.jenvman.2020.110830.
Viveros-Viveros, H., Sáenz-Romero, C., López-Upton, J., & Vargas-Hernández, J. J. (2007). Growth and frost damage variation among Pinus pseudostrobus, P. montezumae and P. hartwegii tested in Michoacán, México. Forest Ecology and Management, 253(1–3), 81–88. https://doi.org/10.1016/j.foreco.2007.07.005
Von Wallis, A., Aquino, D., Knebel, O., y Haberle, T. (2020). Fertilización del cultivo de mandioca para obtención de almidón en Misiones, Argentina. XXVII Congreso Argentino de la Ciencia del Suelo, (pp.1–6). Corrientes.

